微生物在地球生物圈中占据主导地位。在地球约46 亿年的演化历史中,微生物生命存在的时间跨度远超其他生物类群。然而,相较于动植物,细菌的化石证据极度匮乏,其早期演化历史长期缺乏清晰认识。细菌何时出现、何时分化、以及它们如何适应不断变化的地球环境,至今仍未建立起可靠而统一的时间框架。尽管如此,微生物代谢活动在深时地球化学记录中留下了可辨识的信号(Knoll et al., 2016; Lyons et al., 2024),为重建其演化历史提供了重要线索。其中,约24.3-23.3 亿年前发生的大氧化事件(Great Oxidation Event,GOE)被视为地球表生环境演化的关键转折点(Warke et al., 2020; Poulton et al., 2021)。现有证据表明,GOE可能由蓝细菌产氧光合作用的演化所驱动(Dismukes et al., 2001),促使大气中自由氧首次实现持续性积累,不仅深刻重塑了地表环境的化学状态,也为复杂生命的出现奠定了基础。然而,一个长期悬而未决的核心问题是:细菌对氧的适应究竟始于何时?有氧代谢能力的获得,究竟是GOE 后环境氧含量上升的被动响应,还是在GOE之前便已经开始逐步演化?
为回答这一关键问题,近日,来自澳大利亚昆士兰大学、匈牙利厄特沃什·罗兰大学、日本东京大学等十几家机构的研究团队合作在Science发表了一项具有里程碑意义的研究成果(Davín et al., 2025)。该研究整合了细菌基因组数据、机器学习预测、系统发育协调分析和分子钟定年方法,系统探究了地球早期细菌从厌氧到有氧代谢生活方式的转变。通过将GOE与细菌有氧谱系的出现时间相关联,并引入多项新的最大年龄校准约束,在细菌系统发育树上精确映射了从厌氧到有氧生活方式的演化。这项工作首次构建了细菌演化与氧适应的地质时间框架,为理解生命演化与地球氧化历史之间的耦合关联提供了关键证据。
从现代基因组中追溯远古代谢能力
研究团队收集并分析了来自 1007 个现存细菌基因组的数据,涵盖了基因组分类数据库(Genome Taxonomy Database, GTDB)中几乎所有主要的细菌目。在此基础上,他们构建了一棵高分辨率的细菌系统发育树,并将其作为重建祖先性状的骨架(图1)。研究根据 BacDive数据库的标准,将有氧生物界定为能够在有氧环境下生长的类群。该范畴既涵盖将氧气用于呼吸链代谢的生物,也包括仅能耐受氧气存在的生物。研究使用机器学习算法给予基因组的基因组成预测氧适应性,经比对发现,基于梯度提升算法的 XGBoost 模型在BacDive 注释数据的验证下表现最为准确且稳健。该模型以与呼吸链、能量代谢和抗氧化相关的基因组合为特征,对每个细菌基因组进行分类预测。随后,这一预测结果被映射回系统发育树,用于推断祖先节点的代谢状态。
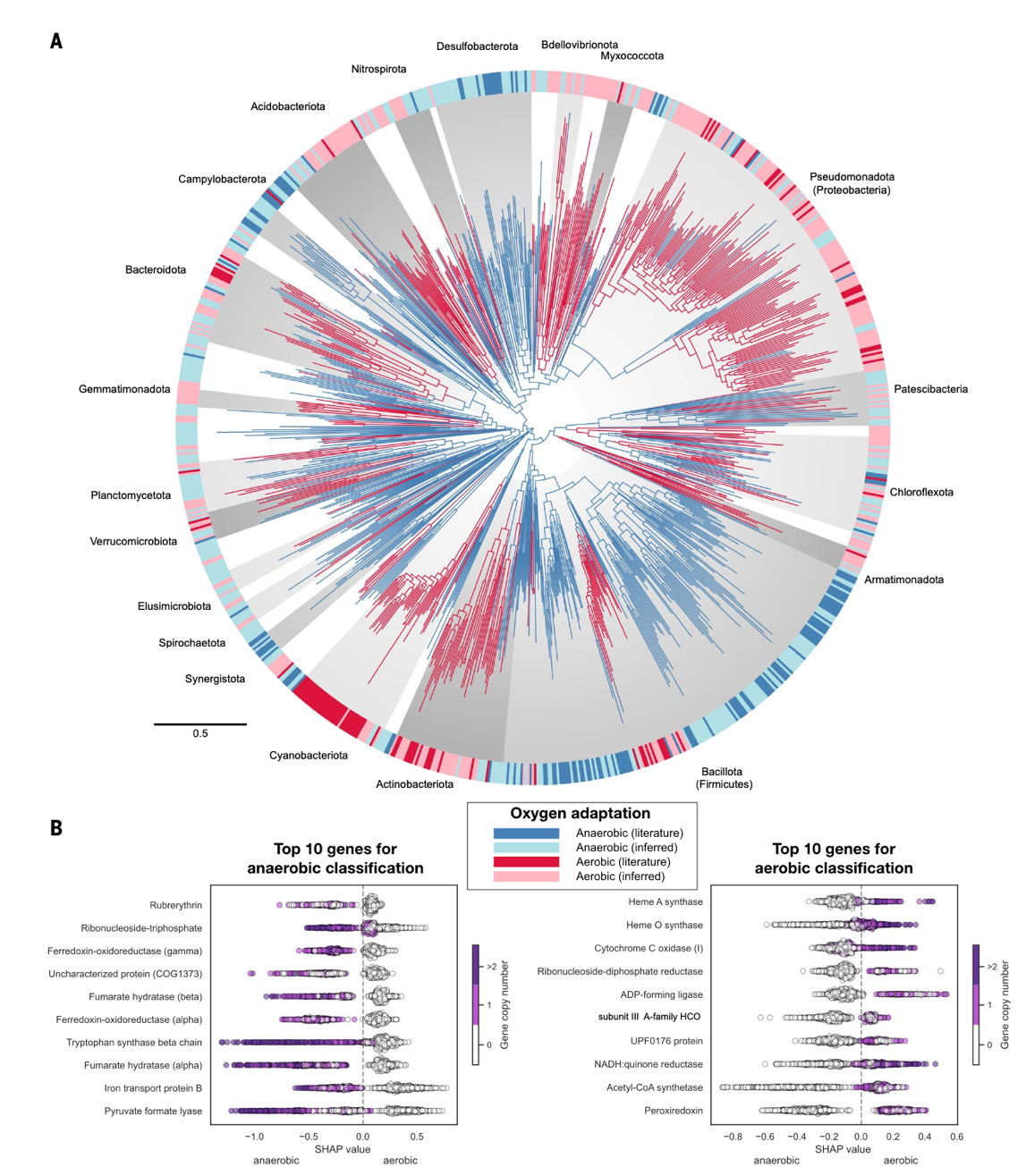

图1 细菌对氧气适应的演化历程。细菌系统发育树(基于1007 个基因组的65 个标记基因),分支颜色表示预测的氧适应能力(红色:有氧;蓝色:厌氧)。外圈显示现存基因组的观测(深色)或预测(浅色)氧适应表型
用GOE事件“校准”细菌演化时间
由于缺乏可直接用于定年的化石证据,细菌分子钟研究长期面临时间校准约束不足的问题。该研究采用了一种创新策略:将 GOE 作为有氧细菌谱系出现时间的“软最大年龄”约束条件,即假定稳定的有氧代谢不可能在大气持续富氧之前大规模出现。同时,研究还引入了与真核生物细胞器的基因相关的化石校准点,包括线粒体(关联α-变形菌Alphaproteobacteria分支)和质体(关联蓝细菌Cyanobacteria分支),从而增加了可用于细菌物种树定年的化石校准信息,使细菌与真核生物的演化时间框架能够在同一时间轴上实现协同校准(图2)。这种多重校准策略显著提高了分子钟估计的稳定性与可信度。
贝叶斯松弛分子钟定年分析结果表明, GOE 软最大年龄约束的引入,使有氧细菌谱系的起源时间估算整体向现在偏移,当应用该校准时,细菌门的平均年龄变年轻约 3.28 亿年。需要强调的是,由于序列数据和校准信息可以克服先验,所以这一结果并非由研究者对 GOE 设定软最大值的假设预先决定。这一发现表明,重大地质事件可为缺乏化石记录的生命类群提供有效的时间锚点。
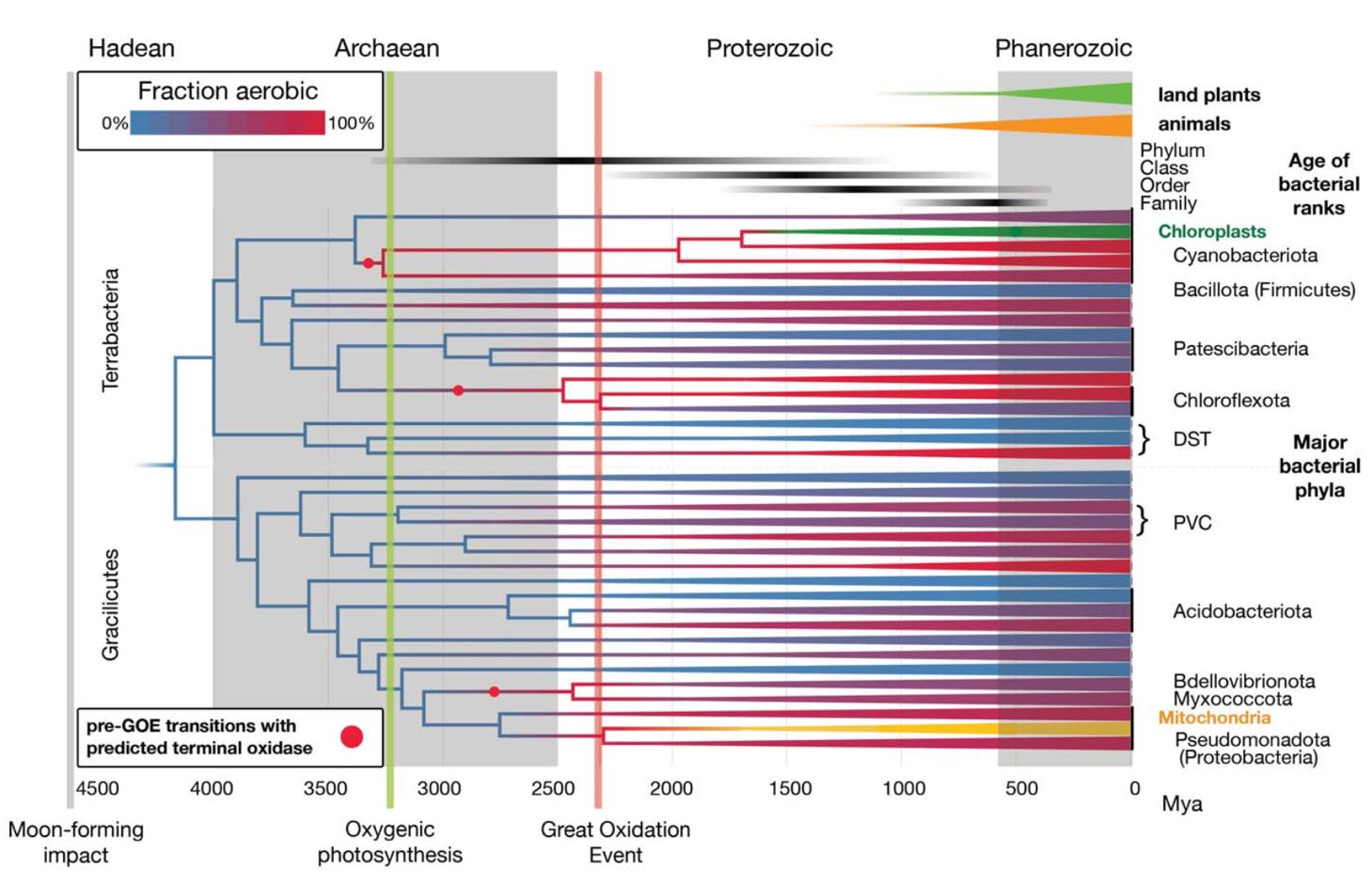
图2 一种用于细菌演化定年与氧适应历史重建的整合分析方法。该方法综合基因组数据、化石记录和地球化学数据,并将细菌的氧耐受性与有氧代谢特征和大氧化事件(GOE)进行关联,从而推断细菌的时间标定系统发育树。图中蓝色表示厌氧状态、红色表示有氧状态;紫色深浅反映了现存细菌门中有氧谱系所占的比例。为充分利用更为丰富的真核生物化石记录,分析中引入了线粒体和质体谱系。同时标示了陆生植物和动物,以便在同一时间尺度下进行对比
重新认识早期生命演化的时间尺度
基于分子钟分析估计出细菌最后共同祖先(Last Bacterial Common Ancestor, LBCA)可能生存于冥古宙至最早的太古宙(约44-39亿年前)。这一时间点晚于月球形成大撞击事件(Moon-forming Impact, MFI),但很可能早于晚期重轰炸期(Last Heavy Bombardment, LHB; 约40-38 亿年前)(图3)。这意味着,在地球形成后不久,生命便已出现并开始快速分化。研究进一步显示,许多现代细菌科在太古宙晚期至元古宙早期(约 25-18 亿年前)就已经形成(图2),演化历史极为悠久。现存线粒体(代表现存真核生物)的最近共同祖先生活于约 18.80-15.02 亿年前。从时间深度来看,真核生物域的整体演化跨度仅大致相当于细菌的一个“纲”。相比之下,后生动物(8.33-6.50 亿年前)和陆生植物(5.15-4.94 亿年前)的冠群年龄,在该研究框架下则更接近细菌“科”级类群的时间尺度(约 8.99-4.57 亿年前,图 2)。
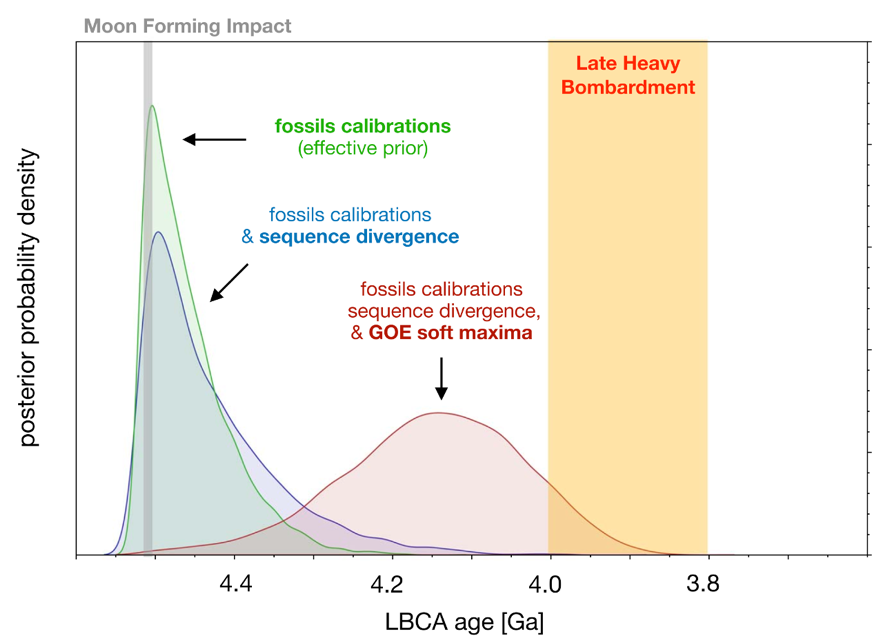
图3 不同分析条件下推断的细菌最后共同祖先(LBCA)年龄。LBCA 的推断年龄相对于月球形成大撞击事件(MFI)以及晚期重轰炸期(LBH)的时间关系。仅基于系统树拓扑结构和定年校准信息(以MFI设定LBCA根节点最大年龄的有效先验,绿色)时,概率主要集中在约45.2 亿年前。加入序列数据后,LBCA 年龄整体偏向更年轻,但仍紧邻MFI时间边界(蓝色)。在此基础上引入GOE软最大年龄约束(红色),进一步减弱了仅由拓扑和校准所导致的过早年龄估计
有氧代谢并非“大氧化事件”的简单产物
祖先状态重建表明,大多数细菌门(38/49)在其起源阶段呈厌氧特性(图2)。该研究在包含1007 个基因组的细菌系统发育树上识别出 84 次从厌氧向好氧生活方式的演化转变,其中绝大多数(72/84)发生于GOE之后。研究中一个尤为引人注目的发现是:并非所有有氧代谢的出现都发生在 GOE 之后。在若干细菌谱系中,研究者识别出发生在 GOE 之前的有氧适应事件。即使在应用 GOE 软最大年龄的条件下,仍有12 个有氧代谢转变节点的后验中位年龄估算早于 GOE,其中 5 个节点的后验概率达到或超过 95%(图 4A),这为细菌在大氧化事件之前已具备氧气适应能力提供了证据。这表明在GOE之前,微生物可能已暴露于局部富氧环境中。这些氧气可能源自非生物过程,例如水的光解或放射分解、硅酸盐岩破碎与水的化学反应等,也可能源自微生物席或细菌早期产氧光合作用出现后形成的海洋氧气绿洲。
最早的一次有氧代谢转变推断发生在约32亿年前,即至少早于GOE约9亿年,出现在蓝细菌纲 Cyanobacteriia 与 Vampirovibrionia 的共同祖先(以下简称 CV 祖先,图4B节点2)。有意思的是,关键的产氧光合作用基因最早出现在蓝细菌纲Cyanobacteriia的祖先中(图 4B节点 3),其时间晚于该谱系中有氧代谢能力的出现。这支持了这样一个假设:有氧代谢能力的获得可能是产氧光合作用演化的必要前提,并可能促进了其进化,因为早期蓝细菌能够利用这种适应性来应对或甚至利用其自身产生的氧气。
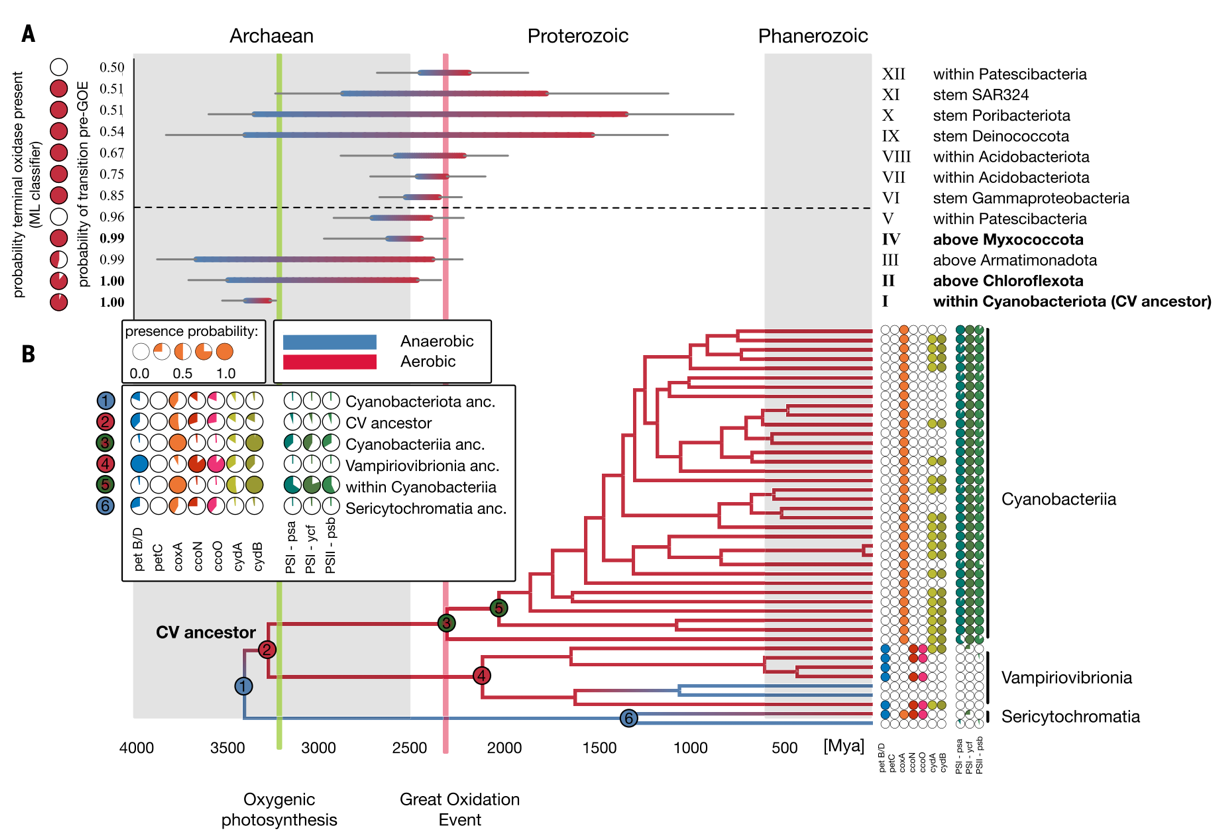
图4 有氧代谢早于大氧化事件(GOE)出现,并可能促进产氧光合作用。(A) 推断的有氧转变年龄范围(蓝色→红色),可能发生在GOE 之前;(B) 蓝细菌门(Cyanobacteriota)中有氧转变及基因演化。推断有氧生活方式的转变发生在CV祖先的分支(节点2),而产氧光合作用出现得更晚,位于Cyanobacteriia纲冠群的干系分支(节点3)。绿色线约3.23 Ga 表示产氧光合作用化石与同位素证据;红色线约2.32 Ga 表示GOE 结束
细菌在地质时间尺度上的演化
通过分析现代与祖先细菌基因组差异随时间演化的模式(图 5A)可以看出,大多数细菌基因组的多样性在约 35 亿年前已基本形成,主要出现在 Terrabacteria 和 Gracilicutes 两大门类的辐射时期,这一结果支持了“太古宙基因家族扩张”(Archaean expansion)的观点。进一步基于物种形成事件间隔时间的指标分析(图5B)表明,有氧适应主要经历两个阶段:首先,自GOE开始,有氧谱系迅速获得分化优势,并在随后约 15 亿年间维持较高水平;之后该优势略有下降,但在过去约 5 亿年中再次回升。此外,通过衡量有氧与厌氧代谢之间转变方向性的指标(图 5C)也显示,有氧谱系具有较高的分化速率。研究者推测,这一演化模式可能源于多种因素的共同作用:GOE的发生创造了大量新的有氧生态位;古生代大气中自由氧水平的第二次持续上升,为有氧谱系提供了更多生态机会;于此同时,部分早期厌氧谱系发生选择性灭绝,其生态位被压缩和分割。在中元古代期间,有氧谱系数量缓慢增加(图 5D),这与当时相对低的大气氧水平一致,这可能限制了有氧谱系的快速扩张。
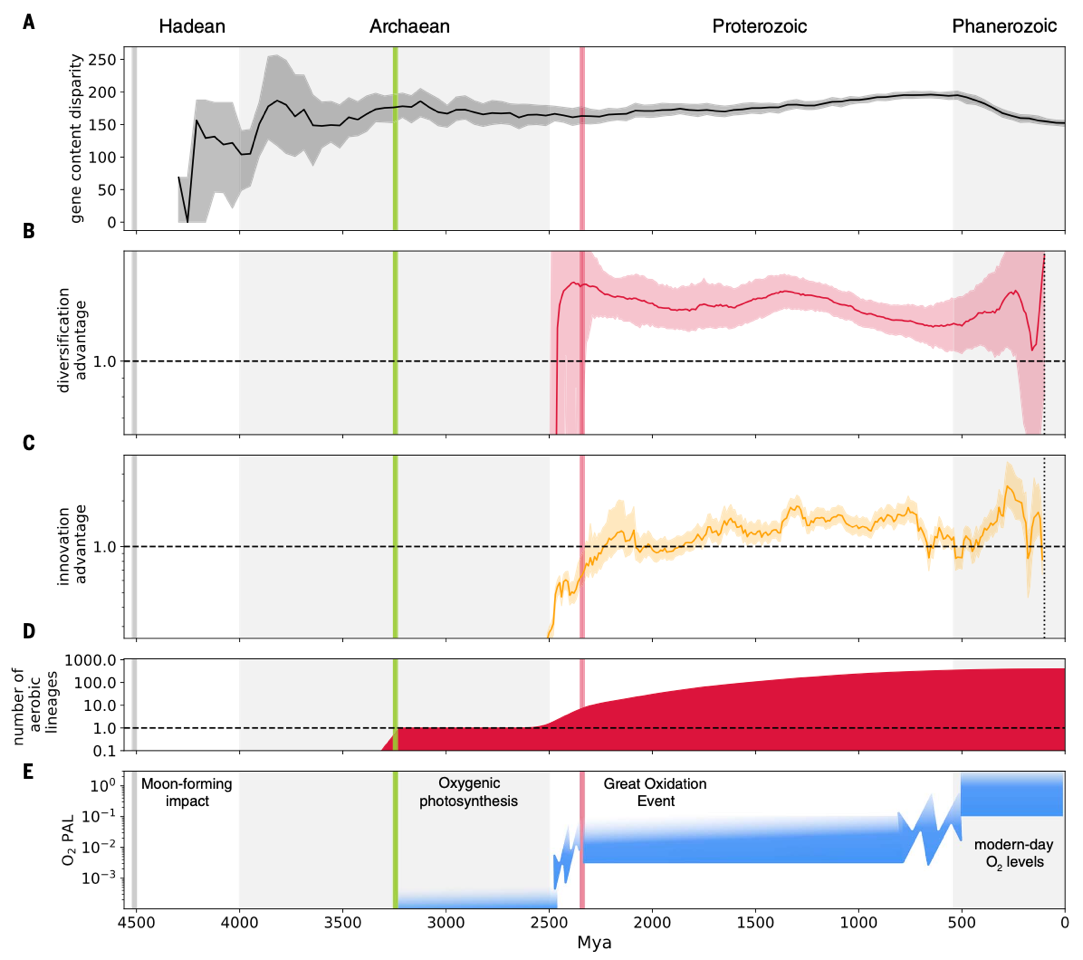
图5 细菌在地质时间尺度上的演化。(A)细菌基因组成差异变化与时间的对应图。(B)曲线显示有氧与厌氧谱系的相对分化速率。(C)水平基因转移推动有氧代谢的扩散。自GOE 以来,大多数时间内从厌氧向有氧的转变次数多于逆向转变。(D)根据图2 显示的有氧谱系数量。(E)大气氧水平随时间的变化示意。灰线表示约4.52 Ga 的MFI;绿线(约3.23 Ga)反映产氧光合作用的化石及同位素证据;红线(约2.33 Ga)表示GOE结束
意义与展望
该研究首次将细菌基因组数据、代谢表型预测与地质事件校准相整合,构建了高分辨率的细菌演化时间框架。研究结果表明,细菌有氧代谢的起源可能早于产氧光合作用和大氧化事件(GOE)。GOE显著促进了有氧谱系的快速扩散与高度分化,且该过程伴随着氧呼吸复合体基因的广泛水平基因转移。该研究不仅深化了对早期生命演化节奏的理解,也为探索生命与地球环境的深时协同演化提供新范式。未来,类似方法可用于解析细菌其他表型的演化,并结合地质历史记录研究古菌域或整个生命域的氧适应过程,从而构建早期地球生物对大气氧化响应的全景图,进一步丰富对生物与地质演化互动机制的认知。
主要参考文献
Dismukes G C, Klimov V V, Baranov S V, et al. The origin of atmospheric oxygen on Earth: the innovation of oxygenic photosynthesis[J]. Proceedings of the National Academy of Sciences, 2001, 98(5): 2170-2175.
Davín A A, Woodcroft B J, Soo R M, et al. A geological timescale for bacterial evolution and oxygen adaptation[J]. Science, 2025, 388(6742): eadp1853.(原文链接)
Knoll A H, Bergmann K D, Strauss J V. Life: the first two billion years[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2016, 371(1707): 20150493.
Lyons T W, Tino C J, Fournier G P, et al. Co‐evolution of early Earth environments and microbial life[J]. Nature Reviews Microbiology, 2024, 22(9): 572-586.
Poulton S W, Bekker A, Cumming V M, et al. A 200-million-year delay in permanent atmospheric oxygenation[J]. Nature, 2021, 592(7853): 232-236.
Warke M R, Di Rocco T, Zerkle A L, et al. The great oxidation event preceded a paleoproterozoic “snowball Earth”[J]. PNAS, 2020, 117(24): 13314-13320.
(撰稿:万娟、张慧卿/地球与行星物理学科中心)