OAI在自然界硫循环中作用巨大。通常而言,OAI下部无氧环境中的硫化氢,需要被带到OAI上部的有氧环境下,才能被彻底氧化成硫酸盐。前人研究发现,微生物演化出多种策略,充当硫物质的“转运使者”或硫氧化-还原反应的电子“传递体”,并从中获得能量,供自己生长。比如,电缆细菌以头尾相接的方式组成一条上千个细胞长度的丝状结构,一头伸出沉积物连接氧气,一头扎入沉积物中吸收硫化物。在下部将硫化氢氧化成单质硫,同时释放电子,再通过身体上的纳米电缆把电子传递到上端,进行有氧呼吸,产生能量,以供细胞生长。与电缆细菌不同,纳米比亚嗜硫珠菌(迄今发现尺寸最大的单细胞细菌,可达750微米)在厌氧环境利用体内存储的硝酸盐将硫化氢氧化成单质硫(S2-→S0),并以硫颗粒的形式暂时存贮起来(在光学显微镜下硫颗粒会发出闪烁夺目的光彩,使得整个纳米比亚嗜硫珠菌细胞泛着微微的“珠光”,像极了饱满圆润的珍珠,因而得名)。当受到水流或其他物理扰动,携带有大量硫颗粒的细胞被带到有氧环境后,会利用氧气将单质硫彻底氧化成硫酸盐(S0→SO42-),同时吸收并将大量的硝酸盐存贮在囊泡中,以备无氧环境下氧化硫化氢所需(图1)。
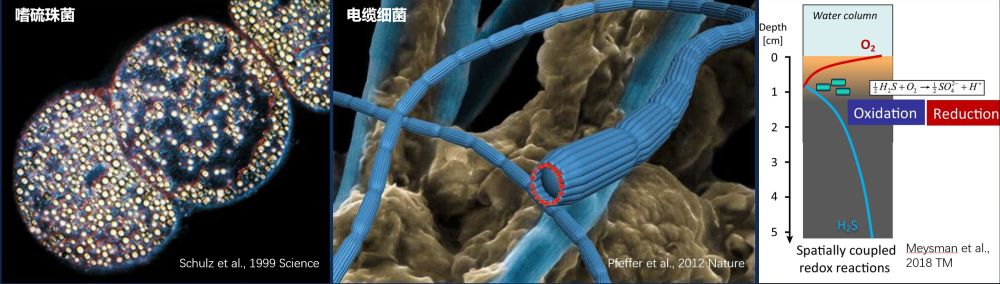
图1 各种硫细菌各显神通跨越“有氧-无氧界面”。电缆细菌充当硫氧化还原的电子传递体,而嗜硫珠菌充当硫物质的转运使者
趋磁细菌是典型的梯度微生物,它们在全球水生环境中广泛分布,且集中生活在OAI界面或稍靠下的厌氧环境中。趋磁细菌能在细胞内合成纳米磁性晶体颗粒(也被称为磁小体,化学成分为磁铁矿(Fe3O4)或胶黄铁矿(Fe3S4)),磁小体多呈链状排列,作为细胞的小磁针,可以感知地磁场,使其沿磁力线方向游泳,从而迅速找到最适合生存的微环境,即水体中“有氧-无氧界面(OAI)”。学界把这种行为称作“趋磁性”或“磁辅助-趋化性”。大量观测发现,趋磁细菌除了在细胞内矿化合成磁小体外,很多还可以在细胞内合成硫、多聚偏磷酸、脂质体、甚至碳酸钙等颗粒,指示趋磁细菌具有C、N、P、S和Fe等元素多样化的代谢潜能,及其在推动这些元素的地球化学循环中的潜在贡献。
为了认识趋磁大杆菌 “趋磁性、生物矿化、生理代谢”三者之间的内在联系,中科院地质与地球物理研究所地磁场与生物圈演化学科组研究团队联合加拿大同步辐射光源实验室和澳大利亚国立大学的合作者,综合透射电子显微学、同步辐射和单细胞宏基因组学技术对发现自天津于桥水库的趋磁大杆菌进行了深入研究。结果发现:
(1)除了已经发现的磁小体(M)、硫(S)和脂质体(L)颗粒外,趋磁大杆菌还能在细胞内形成第四种微米级别大小的囊泡(V)结构,扫描透射电镜电子能量色散谱(STEM-EDS)和同步辐射X射线吸收谱(STXM-XAS)分析表明,囊泡中不含有机物,可能是一种无机物的贮藏结构。统计分析显示,与所有细胞均含有磁小体不同,约24.7%的细胞只含有硫颗粒,约12.9%的细胞只含有囊泡,其余细胞(~62.4%)即不含硫颗粒,也不含囊泡。这指示硫与囊泡只是细胞内的“临时住户”,二者存在某种“此消彼长”的关系(图2)。
(2)同步辐射扫描透射X射线显微谱学(S L-吸收边的X射线近边结构谱,S L-边STXM-XANES)分析表明,趋磁大杆菌细胞内的硫颗粒并不是通常认为的环状硫(S8)结构,而是线状结构,且朝向颗粒内部,其聚合度增加(e.g., S3+→S5+),这可能指示这些硫颗粒处在从外到内的动态降解或合成过程中。
(3)C K-边的STXM-XANES分析表明,趋磁大杆菌细胞内的有机质及其分布具有显著的非均质性,多糖类物质主要分布在细胞外,为胞外多糖组织;脂类和芳香族类有机质在囊泡部位基本没有分布,而在其他部位特别是磁小体链部位显著存在,这与细胞质、磁小体膜及链有机质骨架相匹配;相比较,蛋白质在囊泡膜上显著分布,指示囊泡可能为富含蛋白酶的活性结构(图3)。
(4)N K-边STXM-XANES分析显示,囊泡与细胞其他部位的蛋白质种类及组成明显不同,且能检测到微弱的硝酸盐信号。同时,基因组分析显示趋磁大杆菌不含有合成气泡的任何关键基因,但是具有合成液泡的关键基因,且存在一整套膜结合的硝酸盐还原酶基因。这些实验观察和基因证据均指示,趋磁大杆菌合成的囊泡是液泡,可能用来临时贮藏过多的硝酸盐。

图2 扫描透射电镜电子能量色散谱(STEM-EDS)和同步辐射X射线吸收谱(STXM-XAS)研究趋磁大杆菌细胞结构和化学组成

图3 同步辐射扫描透射X-射线显微谱学技术(STXM-XANES)研究趋磁大杆菌细胞有机质种类及其分布特征
综合前人的研究结果,他们提出了趋磁大杆菌利用地磁场上下穿梭驱动硫循环的新模型(图4)。(1)在OAI下部的无氧环境中,趋磁大杆菌利用硝酸盐将硫化氢氧化成单质硫以颗粒的形式沉积在细胞内(S2-→S0);(2)为了继续完成硫的氧化,细菌需要向上游泳到OAI上部的有氧环境中;(3)在有氧环境中,细胞内存储的硫被彻底氧化成硫酸盐(S0→SO32-→SO42-),同时吸收硝酸根并将其存储在液泡中;(4)怀揣硝酸盐的细菌再次向下游泳到OAI下部的无氧环境,为硫化氢的氧化提供氧化剂。由于北半球地磁场的磁力线是倾斜向下的,因此,在整个过程中,地磁场的定向作用可以把趋磁大杆菌的游动限定在一个近似“上下穿梭”的二维空间中,从而既提高了细菌的穿梭效率,又利于节省能量。另一方面,通过这种上下穿梭,趋磁大杆菌可以完成Fe2+和Fe3+的吸收及其氧化还原,从而合成大量混合价态的四氧化铁颗粒。数目众多的磁小体颗粒也为趋磁大杆菌克服局部扰动或沉积物颗粒阻挠,高效沿地磁场方向定向游泳提供保障。
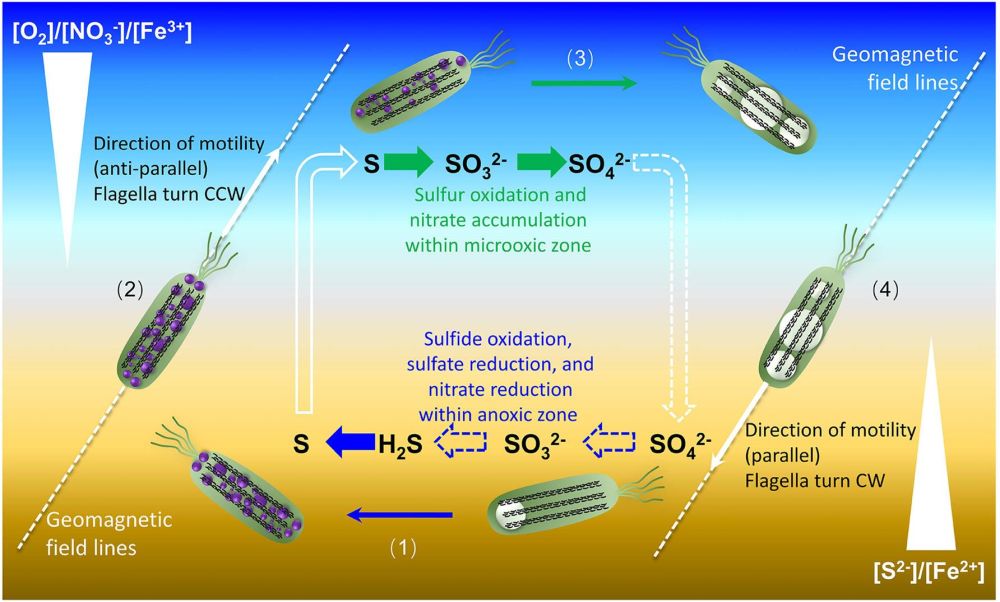
图4 趋磁大杆菌沿地磁场上下穿梭驱动硫循环模式图
其它趋磁细菌有没有可能采用同样的机制呢?他们通过基因组学对比分析了84种代表性趋磁细菌和硫细菌对碳、氮、硫和铁等元素的代谢潜能(图5)。基因预测分析表明,与其他硫细菌相比,趋磁细菌具有更加完善和全面的Fe2+/Fe3+吸收途径,具有成套的磁小体矿化合成相关基因;具有完整的硝酸盐还原和亚硝酸盐氧化酶基因;具有完整的硫氧化(S2-→S0→SO32-→SO42-)和硫酸盐还原(SO42-→SO32-→S2-)关键酶基因。除了δ-变形菌纲趋磁细菌外,其他类群的趋磁细菌均具有碳固定的自养生长和有氧呼吸的异养生长相关的关键酶基因。这指示,趋磁细菌可能采用与趋磁大杆菌相似的策略,利用地磁场的定向作用,在水生环境的无氧环境和有氧环境中上下穿梭,从而驱动OAI中的碳、氮、硫、磷和铁等元素循环。这项研究揭示了趋磁细菌可以利用地磁场的定向作用,在水体有氧-无氧界面上下高效穿梭,从而驱动OAI环境中的物质和能量循环,既指示趋磁细菌是自然环境中驱动碳、氮、磷、硫、氧和铁等重要元素地球生物化学循环的重要微生物功能群,又表明趋磁细菌利用这种高效的穿梭机制,将“趋磁性、生物矿化、生理代谢”三者有机的联系起来,从而使自己能很好地适应厌氧和微氧环境,并从中获取细胞生长和磁小体合成所需要的能量和物质。
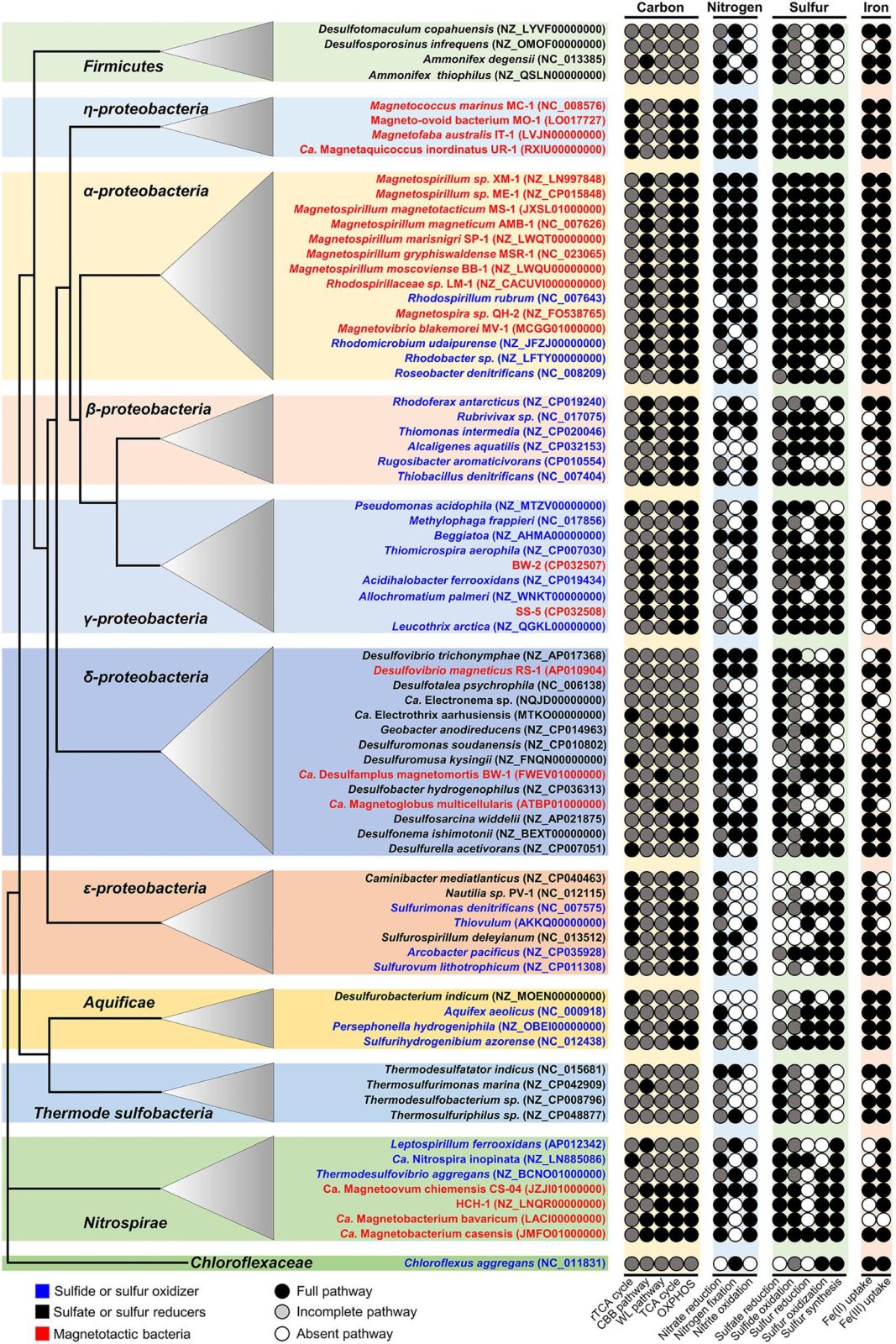
图5 基因预测分析揭示趋磁细菌具有碳、氮、硫和铁等元素多样化的代谢潜能,指示其是一类重要的微生物功能群,在地球水生环境的物质和能量循环中起重要作用
相关研究成果发表在JGR-Biogeoscience上,并被选为AGU亮点文章,并在其EOS网站进行了相关报道。(Li J, Liu P, Wang J, et al. Magnetotaxis as an Adaptation to Enable Bacterial Shuttling of Microbial Sulfur and Sulfur Cycling Across Aquatic Oxic‐Anoxic Interfaces[J]. Journal of Geophysical Research: Biogeosciences, 2020, 125(12): e2020JG006012. DOI: 10.1029/2020JG006012)(原文链接)本项目研究受中国国家自然科学基金重点国际(地区)合作研究项(41920104009)、国家自然科学基金重大项目课题(41890843)和国家自然科学基金创新研究群体项目(41621004)资助。
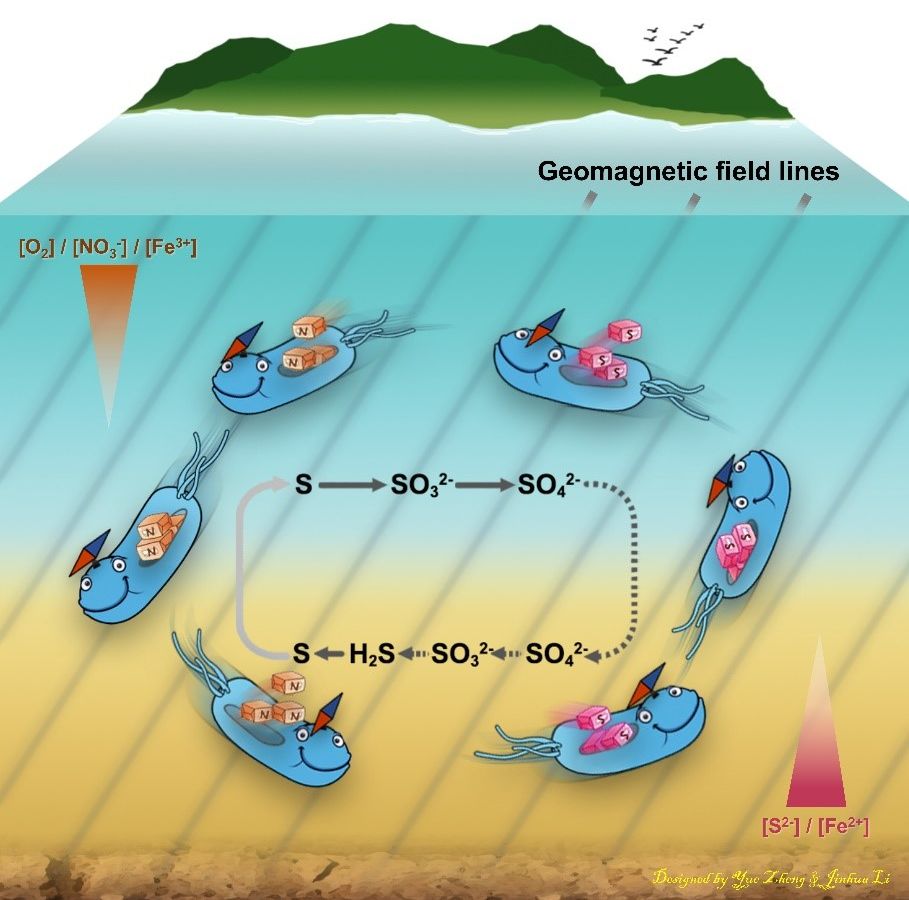
这幅卡通图展示了趋磁细菌利用地磁场的定向作用,在水体中向上向下来回穿梭,不仅将不同类型化学物质上下运输,还将其在OAI上部的有氧代谢过程(硫氧化和硝酸盐吸收等)与OAI下部的厌氧代谢过程(硫酸盐还原、硫的部分还原及沉积和硝酸盐还原等)联动起来,从而促进细胞的生长和磁小体的生物矿化(设计:郑越,李金华)