信鸽、知更鸟、大马哈鱼、海龟、蜜蜂等高等动物可以感知并利用地磁场的导向作用,进行长距离的迁徙或洄游等。高等动物(如脊椎动物)生物矿化形成骨骼和牙齿,具有支撑和防御功能,也是地质记录开展生命演化的重要化石材料。尺寸更小、结构相对简单的微生物能不能感知地磁场?它们在地质记录中能否形成特征性化石呢?
1963年意大利学者Bellini首次在淡水中发现了磁敏感细菌,但他将研究结果以意大利文的形式发表在其所在单位的内部刊物上,未引起大家的关注。1975年,一位美国微生物学博士,在研究海底污泥中螺旋菌时,意外发现了这一类能够沿外加磁场游泳,在细胞内合成链状排列的纳米级四氧化三铁晶体颗粒的微生物,将其命名为趋磁细菌,引起了不同领域科学家极大的兴趣,从此拉开了趋磁细菌研究的序幕。1986年,美国和德国两个研究小组在海洋沉积物中发现了趋磁细菌死亡后遗留的磁性纳米化石-磁小体化石,直接促使地球科学家(特别是古地磁学和海洋磁学领域研究者)加入到趋磁细菌和磁小体化石的研究中。1996年美国NASA科学家宣称,他们在著名的火星陨石ALH84001中发现了疑似磁小体化石的纳米级磁铁矿颗粒,再次掀起了从古老沉积物和岩石中寻找磁小体化石开展早期生命、古环境和古地磁研究的热潮。
经过了四十多年的研究,来自生命科学、地球科学和材料科学等多个领域的研究者,在趋磁细菌的生态分布和种类多样性、磁小体生物矿化过程及其分子机制、磁小体化石识别及古地磁学应用、以及磁小体生物仿生和纳米医学应用等方面开展了大量研究。磁小体合成的大致路径和多个相关基因及其功能已经被明确,诸多岩石磁学、矿物学和晶体学参数已经被提出,用来识别磁小体化石。
然而,目前对磁小体生物矿化的精细过程(尤其是晶体生长和磁性变化的无机过程)及其多样性的认识严重不足。从地质记录中识别磁小体化石并利用其开展古地磁、古环境和早期生命研究仍然是一大挑战。主要原因在于:
(1)对磁小体晶体生长机制的初步研究只在少数几种实验室可培养的趋磁细菌中进行,这严重制约了对趋磁细菌磁小体多样性及其生物矿化机制的全面认识;
(2)基于少数几种趋磁细菌研究而建立的磁小体化石识别标准和方法存在很大的局限性,甚至有些标准并不准确,这制约了寻找识别磁小体化石的进度,完善原有的识别标准并建立新的磁小体化石识别标准迫在眉睫;
(3)缺乏系统的微生物分子生态学、电子显微学和纳米磁学研究,导致“趋磁细菌种类-磁小体生物矿化-磁小体磁学性质”三者的内在联系还没建立,造成我们目前还不能从磁小体化石的磁学性质和晶体结构特征获得地质历史时期趋磁细菌的生物学和生态学信息,这极大地限制了磁小体化石的古地磁学和古环境学应用。
针对这些问题,近五年来中科院地质地球物理所地磁场与生物圈演化学科组的研究团队联合中科院物理所、法国巴黎第六大学、澳大利亚国立大学等国内外多个单位的科研人员,对环境中未培养的趋磁细菌展开了生物地磁学和地球生物学多学科交叉研究。他们首先建立了一种“荧光-电子显微镜联用”新技术(FISH-SEM和FISH-TEM)(图1),将趋磁细菌种类鉴定的荧光显微镜观测信号与磁小体结构观测的电子显微镜观测信号结合起来,首次在单细胞水平实现了未培养趋磁细菌的种类鉴定和生物矿化研究。应用该技术,他们已经从北京及周边区域湖泊(如密云水库、北海)、西安及周边区域湖泊(如未央湖、护城河和咸阳湖等)、以及河北秦皇岛近海沿线的半咸水和海洋环境(如汤河、石河入海口和七里海等)中已经发现了四个门类超过二十余种趋磁细菌新菌种。随后,他们应用同步辐射技术,对每一种趋磁细菌的一个细胞开展从微纳米尺寸到原子水平的综合研究。
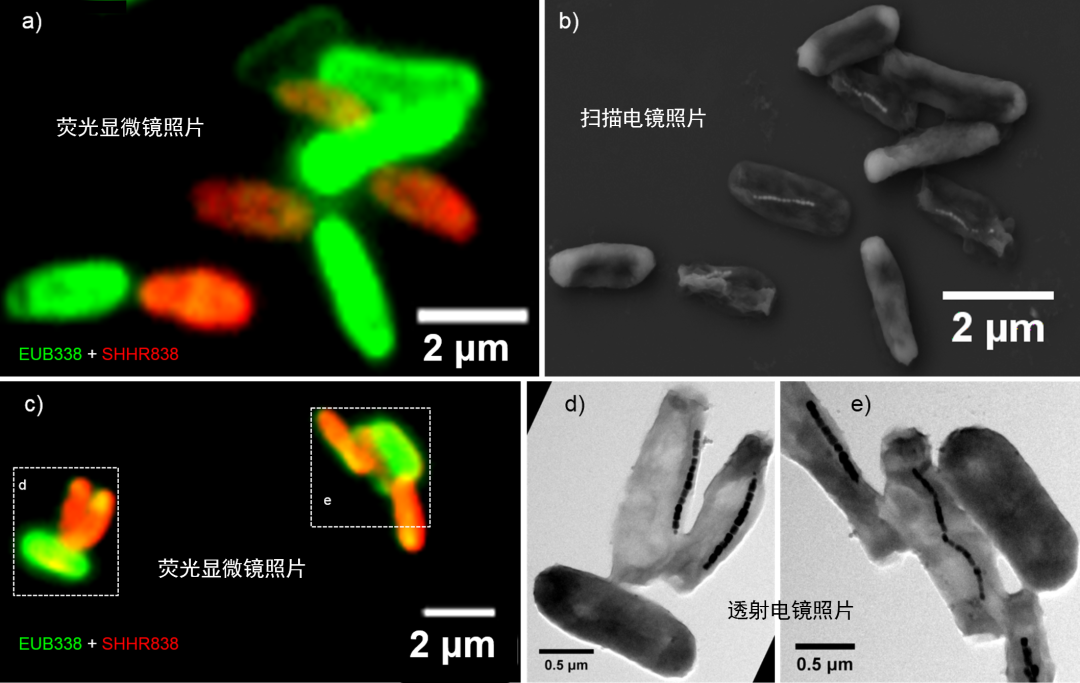
图1 “荧光-电子显微镜联用”新技术(FISH-SEM和FISH-TEM)。该技术基于趋磁细菌的16S rRNA基因序列,设计特异性的寡核甘酸探针,通过改进传统的荧光原位杂交(FISH)技术的制样和观察环节,实现了荧光显微镜下对经寡核苷酸探针特异性杂交的趋磁细菌进行单细胞水平的观察和识别,然后通过扫描电镜和透射电镜对同一微区或同一个细胞进行纳米尺度的形貌和结构分析,从而实现对同一个趋磁细菌细胞在单细胞水平上的种类鉴定(系统发育)和在纳米尺度上的生物矿化研究(细胞和矿物形貌结构等)(Li et al., 2017)
近期,在对来自西安未央湖沉积物中的豆形趋磁性杆菌WYHR-1进行细胞形态、化学成分及其磁小体的晶体生长过程的研究时,他们发现趋磁细菌能在细胞内矿化合成“纳米尺寸大小、晶体结构独特”的磁铁矿(Fe3O4)颗粒(也被称为磁小体),磁小体的晶体生长过程和晶型多样,但在同一类群或同一种趋磁细菌中却具有独特性。
研究发现,WYHR-1在细胞内合成“子弹头形”磁铁矿晶体颗粒,具有典型的“多阶段晶体生长”模式:颗粒初始各向同性生长到~20-22 nm时,开始各向异性生长(长度增加速度大于宽度增加速度)到长度~75 nm和宽度~35 nm后,颗粒宽度基本保持不变,长度增加到~150-180 nm,个别颗粒(~1.1%)长度可达~280 nm。而且,WYHR-1磁小体在初始生长阶段,无明显晶型,各向异性生长时,始终沿[001]方向拉长生长。成熟的WYHR-1磁小体具有标准的子弹头形,颗粒底部相对平整,为一个较大的{100}面,颗粒侧面圆柱形,顶部圆锥形,少数颗粒顶端残留一个小的{100}面。对整个磁小体链进行HAADF-STEM三维重构和STEM旋进电子衍射(即ASTAR)分析,结果表明WYHR-1磁小体链束大致由2-3条磁小体链紧密排列构成,单个颗粒均沿[001]方向排列成链(图2)。

图2 综合透射电镜技术研究趋磁杆菌WYHR-1细胞和磁小体形状和结构特征及其子弹头形磁小体生长机制
将这些研究结果与科研团队前期研究对比发现,WYHR-1与其它趋磁细菌都不同,其磁小体的晶体生长过程及其晶型具有独特性。比如:
- 趋磁球菌SHHC-1磁小体为八面体形磁铁矿晶体(Zhang et al., Front Microbiol, 2017);
- 趋磁螺菌AMB-1磁小体为稍微拉长的立方-八面体形 (Li et al., Geochem Geophys Geosyst, 2013);
- 趋磁杆菌SHHR-1磁小体为拉长截角六棱柱形(Li et al., Appl Environ Microbiol, 2017);
- 趋磁大杆菌MYR-1为拉长的、弯曲的子弹头形,其磁小体也具有“多阶段晶体生长”模式,但初始阶段晶体各项同性生长成为立方-八面体,随后颗粒多沿[112]或[114],甚至[111]方向拉长生长到一定阶段后,改变拉长方向,最终统一沿[001]方向拉长生长,因此,成熟MYR-1磁小体颗粒多为弯曲的,其颗粒底面为一较大的{111}面 (Li et al., J R Soc Interface, 2015)。
综合这些研究,他们提出磁小体的生物矿化模式(至少从晶体生长方面)具有多样性,然而其晶体生长过程在基因层面上受不同趋磁细菌类群或菌种/菌株的严格调控,其晶型因而具有特异性(图3)。趋磁细菌磁小体是潜在磁性纳米化石,在未来的研究中,可以通过分析沉积物中的磁小体化石的形貌特征及其相应的磁性性质,可以获得古趋磁细菌类群或种类的信息,从而同时开展“古趋磁细菌、古环境和古地磁场”研究。
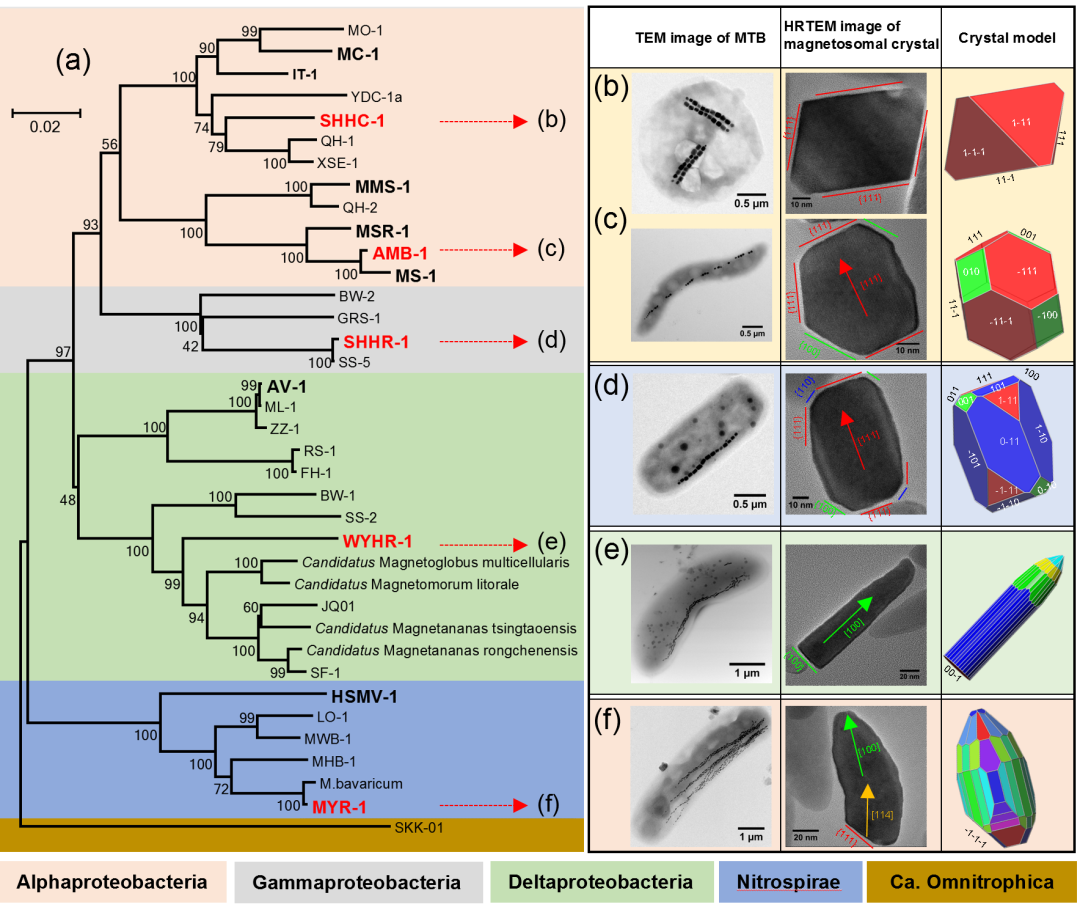
图3 不同类群和种类趋磁细菌的磁小体晶体生长和晶型特征
相关研究成果发表于JGR-Biogeosciences,并被选为封面文章。(Li J, Menguy N, Roberts A P, et al. Bullet‐shaped magnetite biomineralization within a magnetotactic Deltaproteobacterium: Implications for magnetofossil identification[J]. Journal of Geophysical Research: Biogeosciences, 2020, 125(7): e2020JG005680. DOI: 10.1029/2020JG005680)(原文链接)。本项目研究受中国国家自然科学基金重点国际(地区)合作研究项(41920104009)、国家自然科学基金重大项目课题(41890843)和国家自然科学基金创新研究群体项目(41621004)资助。

封面上的趋磁细菌图展示了5种不同类型的趋磁细菌。从系统发育上,它们分属两大门类:变形菌门(上部四个)和硝化螺菌门(右下大杆菌,分离自北京密云水库);上部的四个趋磁细菌又进一步分属于δ-变形菌纲(左上豆形杆菌,分离自西安未央湖),γ-变形菌纲(右上杆菌,分离自秦皇岛石河入海口),η-变形菌纲(中间球菌,分离自秦皇岛石河入海口),和α-变形菌纲(左下螺旋菌,为研究的模式菌株,上世纪九十年代初分离自日本一淡水湖泊)(图/中国科学院城市环境研究所郑越博士)