早期火星存在适宜生命起源的各种条件,比如地表和浅表的液态水、干湿交替的气候,以及生命所需的各种化合物(Sasselov et al., 2020)(见前沿报道《SA:比较行星学视角看生命起源》),这些条件都被火星上的沉积岩所记录。火星沉积岩的形成时间普遍早于地球沉积岩,由于近几十亿年来火星缺乏构造运动和地表水,其沉积岩变质程度更低,故火星比地球更有记录和保存早期生命和前生命化学等相关信息的潜力(Sasselov et al., 2020)。火星车和轨道器数据显示,盖尔陨石坑和耶泽罗陨石坑含有大于35亿年前的无定形二氧化硅、黏土矿物和碳酸盐矿物(碳酸镁)等。同样,地球上最早的生命记录也发现于碳酸盐岩或相关的沉积相中(Lepot, 2020; Javaux, 2019; Brasier et al., 2015)。通过类比研究地球太古宙碳酸盐岩、陆相碳酸盐岩、水合碳酸镁矿物、陆相硅质岩以及碳酸盐-黏土矿物组合中的生命信号,有助于理解耶泽罗陨石坑沉积的天体生物学意义,为“毅力号”火星采样返回和原位数据解释提供指导(Bosak et al., 2021)。
地球太古宙碳酸盐岩由于经历重结晶、变质等改造作用,其保存的生命信号需要结合形貌、结构、有机组分和同位素等多种手段来进行识别(Lepot, 2020; Javaux, 2019; Brasier et al., 2015)。研究表明太古宙碳酸盐岩及其相关的硅质岩具有不同的保存生命信号的方式,太古宙碳酸盐岩可以保存有机质和微生物纹层,而硅质岩通过白云石、黄铁矿和铁氧化物等矿化作用还能保存微生物实体化石(表1)。无定形二氧化硅和高镁碳酸盐矿物如原生和早成岩阶段形成的白云石,可以利用细胞或者有机质表面进行成核作用,来保存生物结构和实体化石。无定形二氧化硅和白云石可以同时出现,但微生物精细结构和实体化石通常保存于最早成岩时期形成的纳米级无定形二氧化硅颗粒中。金属离子可以在二氧化硅浓度非过饱状态和非热液环境中调控微生物硅化作用,例如铁离子能够充当绑定细胞表面和硅元素之间的桥梁,因此厌氧富铁微生物群落容易发生硅化作用。此外,氧化条件下镁离子也有利于微生物硅化,比如形成无定形富镁二氧化硅矿化蓝细菌。富镁燧石和白云石或者铁氧化物和富铁硅质白云石之间的潜在关联,可以为火星富铁镁碳酸盐岩潜在的有机质硅化提供参考模型(Bosak et al., 2021)。
表1 太古宙碳酸盐岩生命信号的研究案例(总结自Bosak et al., 2021)
|
时间 |
地点 |
地层 |
有关生命信号 |
|
35亿年前 |
澳大利亚 |
Dresser组 |
硅质岩-白云岩中毫米宽掌状结构黄铁矿、包含其中的有机质及其同位素信号,指示硅质岩-白云岩早成岩阶段可能已经出现细菌硫酸盐还原作用;
硅质和硫酸钡充填、锐钛矿覆盖的圆形结构,可能是胞外聚合物捕获的热泉水体表面的气泡。 |
|
34.3亿年前 |
澳大利亚,Warrawoona |
|
大型圆锥状叠层石形貌:矿物纹层的弯曲率和前扩散边界层厚度的关系;
非圆锥状叠层石的纹理;
穹窿状叠层石和碳酸盐矿物含有有机薄层;
生物成因碳酸盐岩的形貌、大小、成分和产状。 |
|
34亿年前 |
南非Barberton Greenstone Belt |
|
硅质岩中的微生物结构和化石;
有机碳、硫、氮和氙同位素指示有机质与围岩同生,可能是氢细菌。 |
|
32亿年前 |
南非 |
Moodies群 |
砂岩中燧石和白云石保存的微生物席和碳质纹层;
硅化的丝状结构和微生物形成的气体。 |
|
30亿年前 |
南非 |
Chobeni组 |
叠层石中包裹的颗粒状或薄层状石英、黏土等碎屑矿物出现的角度大于休止角,指示粘性有机质如生物膜的存在。 |
|
28亿年前 |
加拿大 |
Steep Rock组 |
叠层石中包裹的颗粒状或薄层状石英、黏土等碎屑矿物出现的角度大于休止角,指示粘性有机质如生物膜的存在。 |
|
27亿年前 |
南非Hartbeesfortein盆地 |
Ventersdorp超群 |
湖相叠层石中硅化的围绕气泡的透镜体结构和细丝结构,可能是产氧光合作用。 |
火星碳酸盐岩属于非海相碳酸盐岩沉积,因此可以通过对地球上非海相碳酸盐岩的研究来为火星提供参考模型。地球上非海相碳酸盐岩若低于20-30℃形成叫做冷水型钙华,高于这个温度的则叫做热成因型钙华。碳酸盐岩的形貌和矿物组合反映了其形成过程中水化学、水文、气候、溶液饱和度、物质来源、流体下坡或侧面流动、水-气界面矿物沉积以及微生物活动的情况。钙华具有多孔构造,该构造可以用非生物机制解释。当多个碳酸盐凝块与生长面的角度不能用向下沉淀来解释,就可能是由于生物捕获或者生物膜内沉积形成的。非生物成因碳酸盐岩宏观形貌还可以表现为平坦指状,生长过程碳酸盐薄层不断扩大,但厚度不变,其化学组成和类型不能用于判断是否为生物成因。现代热泉碳酸盐结壳由放射纤维状和栅栏状方解石或者文石晶体组成,其生长轴线垂直于生长表面,为非生物成因,但是,这种碳酸盐壳可埋藏生物碎屑。
相比于非生物成因的碳酸盐岩沉积,显生宙以来栅栏结构的硅华可能是生物成因,例如硅华深色少孔纹层和浅色多孔纹层是由于流体性质和气候变化引起的蓝细菌丝状结构取向变化导致的,记录了生物对气候的响应。然而,扩散限制聚集的非局部生长模型在间歇性环境变化下也会影响晶体的冷却、饱和以及生长速率,通过针尖优先生长,在没有生物的影响下可以形成层状、分枝、多孔的掌状结构。蒸发型热泉中的条带结构和与拉长孔隙相关的掌状结构同样没有蓝细菌的参与。因此,非海相碳酸盐岩和硅质岩的生命信号还依赖于其他证据的支撑,比如共生有机质的分布和促进生物硅化的元素(镁、铁、铝)富集(Bosak et al., 2021)。
火星碳酸盐矿物主要为水合碳酸镁矿物,因此在与地球类比时需要考虑两个行星上矿物组合、形成环境/机制以及各自保存生命信号潜力的差异。土耳其Salda湖、澳大利亚昆士兰Gumigil-Kunwarara沉积和英国哥伦比亚盐湖,可以作为最类似火星碳酸镁矿物形成的环境。地球陆相碳酸盐岩以方解石和文石为主要成分,以钙华和岩石涂层的形式分布于地表和浅次表层,由含溶解二氧化碳的表层水与超碱性水反应形成,其δ13Ccarb和δ18Ocarb值很低而且有镉富集现象。根据动力学理论,非生物成因白云石和菱镁矿在低于60-80℃时不利于形成,因此富镁碳酸盐岩在陆相碳酸盐岩中分布较少,但水合碳酸镁矿物可以在较低温度下非海水环境中形成。在超基性地台上,蛇纹石化作用形成的碳酸镁和氢氧化镁主要出现在与大气隔绝的岩脉和裂隙中,其厚度为毫米-分米尺寸,以结壳的形式将蛇纹石化的超铁镁质碎屑、矿脉和矿洞胶结在一起。在Gumigil-Kunwarara超铁镁质高地,碳酸镁以岩脉充填的形式,在沉积低地以细粒胶结物和结核存在。在Salda湖,水合菱镁矿以结壳形式沉淀在蛇纹岩基岩外部和叠层岩堆上端。在哥伦比亚水菱镁矿-菱镁矿干盐滩,蒸发和毛细作用使菱镁矿和水合菱镁矿在水柱或者浅表层形成混合土堆。这些环境都类似于耶泽罗撞击坑碳酸镁沉淀的环境,周边超基性岩石风化带来富镁流体,搬运到沉积盆地内(图1)。如果火星碳酸盐岩分布及其地球化学特征如上所描述,那么其形成过程很可能类似于地球相关沉积。
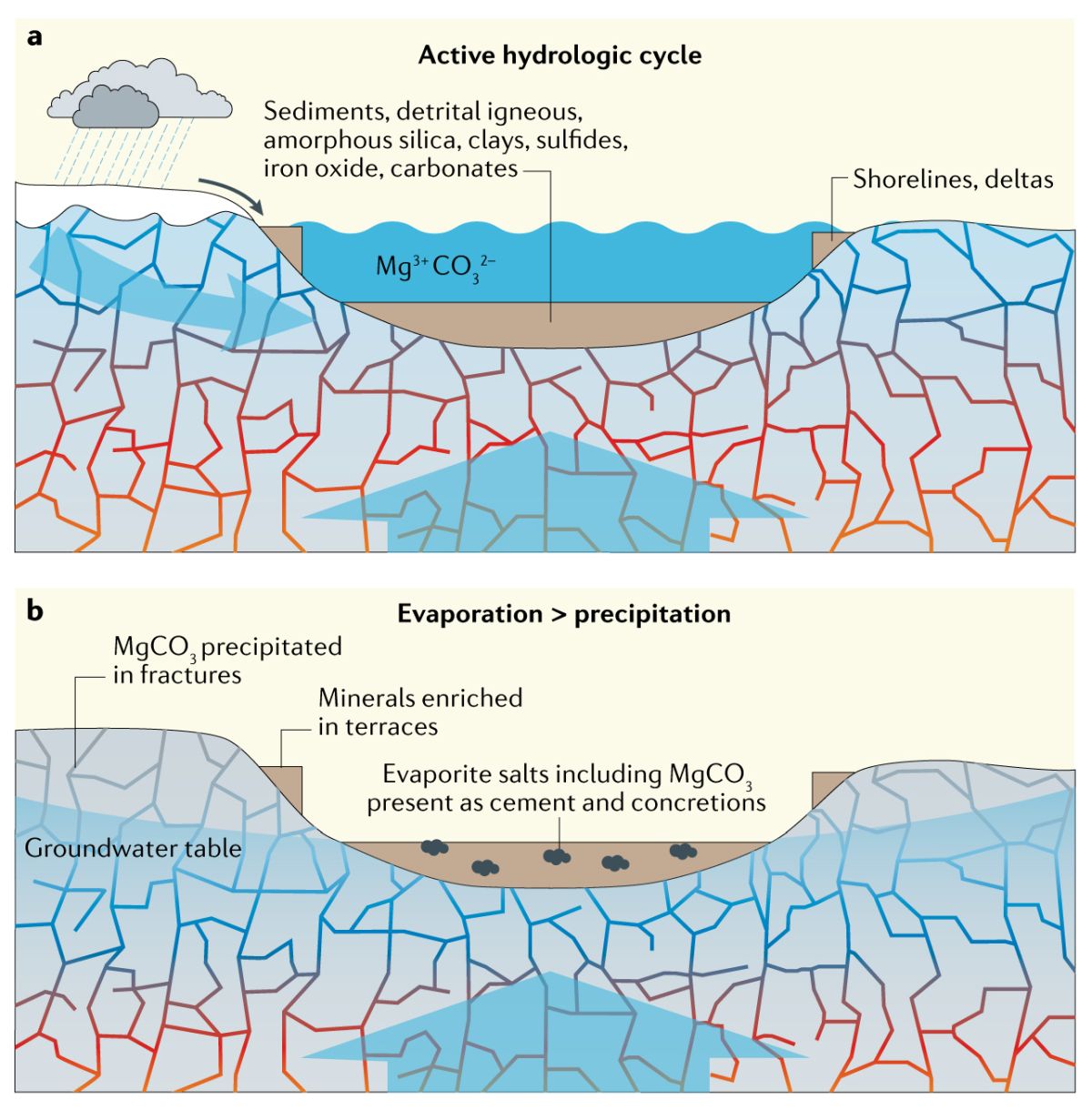
图1 超铁镁质地台中高镁碳酸盐岩的沉积模式(Bosak et al., 2021)
现有研究表明微生物可能参与水合菱镁矿的沉淀,同时水合菱镁矿也有保存生物结构的能力。水合碳酸镁不稳定,容易发生重结晶作用,因此水合碳酸镁保存生物结构和有机质需要特殊的环境。Salda湖和哥伦比亚干盐滩盐壳和叠层石中保存了毫米级别的微生物纹层。Salda湖叠层石和超基性岩覆盖了可能由蓝细菌和硅藻形成的胶质涂层,其中还含有微晶板状的水合碳酸镁矿物,说明微生物活动或者胞外聚合物的降解控制了水合碳酸镁矿物的形成。
另外,超基性岩低温风化流体会富集硅而形成与碳酸盐矿物共生的黏土矿物和水合无定形二氧化硅。早成岩形成的无定形二氧化硅和碳酸盐矿物具有不同的保存化石和生物结构的能力,二氧化硅可以很好地保存蓝细菌结构和部分有机化合物,文石可以保存α变形杆菌。富铁镁质皂石等黏土矿物也有利于保存有机质,保存程度取决于有机分子类型和结构铁的氧化还原状态。陆地上与湖相白云岩共生的原生或者次生黏土矿物如海泡石和皂石可以保存高达3%的有机质,这些黏土矿物以有机质为核心进行成核生长(Bosak et al., 2021)。
对地球类火星沉积或环境的持续研究,包括对影响同位素信号、有机化合物、微生物结构和化石保存过程的研究,将有助于理解火星沉积物的天体生物学意义。研究地球早期生命信号在水合二氧化硅、碳酸盐和自生/碎屑黏土矿物中的保存机制,及其与矿物的共生关系,有助于理解地球早期生命与环境的协同演化以及在火星上寻找潜在的生命(Bosak et al., 2021)。
主要参考文献
Bosak T, Moore K R, Gong J, Grotzinger J P. Searching for biosignatures in sedimentary rocks from early Earth and Mars[J]. Nature Reviews Earth & Environment, 2021, 2: 490–506.(原文链接)
Brasier M D, Antcliffe J, Saunders M, Wacey D. Changing the picture of Earth's earliest fossils (3.5-1.9 Ga) with new approaches and new discoveries[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112: 4859-4864.
Javaux E J. Challenges in evidencing the earliest traces of life[J]. Nature, 2019, 572: 451-460.
Lepot K. Signatures of early microbial life from the Archean (4 to 2.5?Ga) eon[J]. Earth-Science Reviews, 2020: 103296.
Sasselov D D, Grotzinger J P, Sutherland J D. The origin of life as a planetary phenomenon[J]. Science Advances, 2020, 6: eaax3419.
(撰稿:陈妍,申建勋,林巍/地星室)