汇聚板块边缘是两个相互汇聚和消亡板块间的边界,包括一系列重大地质过程:大洋板块俯冲到大陆板块之下,俯冲板块在不同深度发生变质脱水和部分熔融,所产生的流/熔体交代上覆地幔楔导致壳幔相互作用,地幔楔部分熔融引起弧岩浆作用形成弧型新生地壳,新生地壳部分熔融和分异结晶产生陆壳等,因而是连接地球深部圈层与表生层圈的关键区域,也是理解和发展板块构造理论的核心所在(Schellart et al., 2010)。在板块构造理论框架下,将汇聚板块边缘的构造活动与微生物生命过程研究相偶联,不仅有助于阐明深部地质活动对生物圈的影响过程及机制,还能更好地促进固体地球科学与生命科学在整个地球动力系统下的深度交叉融合,从而深入探讨构造活动从宏观到微观尺度上与微生物活动之间的内在联系和协同机制(张传伦等, 2018)。
在最近的一项研究中,美国田纳西大学的Karen G. Lloyd教授和意大利海洋生物资源与生物技术研究院的Donato Giovannelli教授等人合作,将微生物种类及分布与汇聚板块边界的地下挥发性物质输送的变化进行相关性分析,获得了地球深部构造活动与微生物生命过程之间相互作用的直接证据,相关的研究成果发表在Nature Geoscience上(Fullerton et al., 2021)。
哥斯达黎加汇聚板块边缘属于典型的剥蚀型汇聚板块边缘。在该地区,科科斯板块(Cocos Plate,太平洋板块的一部分)以80-90 mm/年的速度向加勒比板块(Caribbean Plate)俯冲,浅俯冲的几何特征促进了俯冲板片脱水,释放出大量的碳和还原性的挥发物进入上覆板块(图1)。该研究涉及的取样位置涵盖了哥斯达黎加汇聚板块边缘的不同俯冲构造单元,包括岛弧区(板片俯冲深度100-120 km,温度>60°C,pH值0-3)、弧前区(俯冲深度40-100 km,温度40-60°C,pH值4-7)和外前弧区(板片俯冲深度20-40 km,温度20-40℃,pH值8-10)(图2)。研究者利用铜管从上述不同构造单元的21个热泉处收集新鲜的流体,并通过流体氦同位素(3He/4He)和溶解无机碳(DIC)的碳同位素(13C)分析证明了这些流体起源于地球深部。最终,他们系统地分析了上述21个热泉流体中的微生物群落组成以及相关的环境参数(温度、比电导率、pH值、DIC与DOC浓度、阴阳离子含量等),从宏观尺度的地球深部构造活动对微生物多样性的影响,以及微观尺度的微生物活动对宏观尺度的地球深部构造活动的反馈这两个方面出发,揭示了深地生物圈与地球深部构造活动之间的联系。
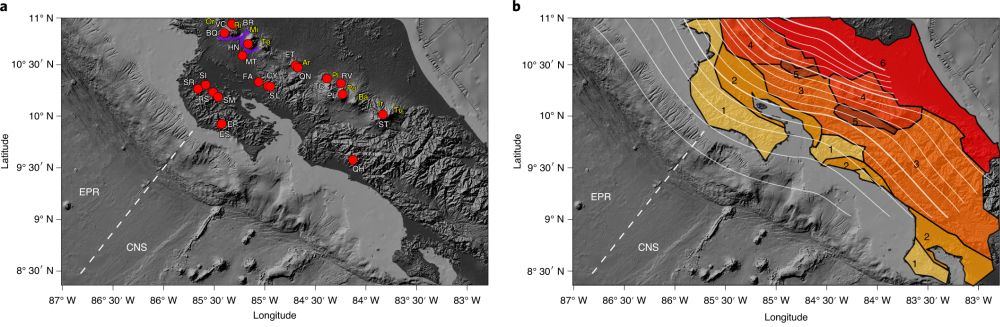
图1 采样点(红色圆圈)在哥斯达黎加汇聚板块边缘的分布情况(Fullerton et al., 2021)。(a)其中白色字体显示的采样点为温泉和火山口湖,黄色字体显示的采样点为火山,紫色线条指示瓜纳卡斯特地热省火山口的位置;(b)俯冲板块深度线和主要岩石类型:(1)白垩纪/第三纪蛇绿岩;(2)第三纪盆地;(3)第三纪火山区;(4)第四纪火山区;(5)弧内盆地;(6)加勒比海岸平原。虚线代表东太平洋中脊(EPR)与科科斯-纳斯卡板块(CNS)的边界
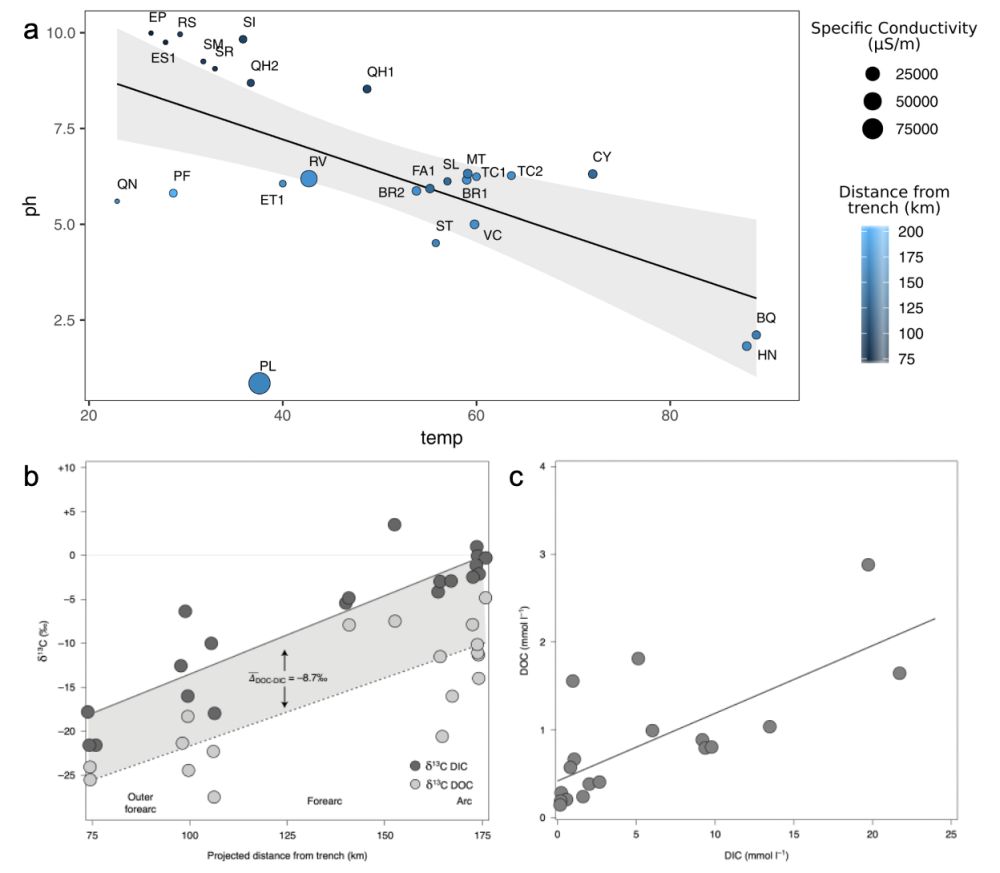
图2 (a)热泉流体pH与温度、比电导率、板块俯冲深度等参数之间的相关性;(b)热泉流体δ13C(DIC和DOC)与板块俯冲深度之间相关性;(c)热泉流体中DIC 浓度与DOC浓度的相关性(Fullerton et al., 2021)
他们利用流式细胞仪与分子进化时钟16S rRNA基因测序技术,对各热泉流体中细菌的细胞丰度及组成类群进行了分析。结果表明,热泉流体中细菌的细胞丰度大致在1.5 ×103 ~ 3.3 ×106 cells/mL,细菌的组成类群也比较丰富,包括了59个分类门。为了全面地认识环境因子对微生物群落结构及分布的影响,该研究开展了多项统计分析。温度和pH环境因子与微生物多样性的关系表明,嗜热微生物(热袍菌门、产水菌门、暗黑菌门)的丰度随温度和pH的升高而增加;相比之下,变形杆菌门和其它一些未培养细菌类群则呈现出相反的趋势(图3)。这说明温度和pH值能够在一定程度上影响和改变微生物的多样性与分布。统计分析还表明汇聚板块边缘微生物的多样性也受到了热泉流体阴阳离子成分的显著影响,而阴阳离子成分则主要受控于与板片俯冲的深度以及形成的深部围岩类型(图4)。进一步的共现网络分析、随机森林模型和关联分析等统计方法发现至少有10个微生物群组(占总微生物类群的99%以上)与不同的地球深部环境因子(DOC、DIC、pH、铁元素、溶解氧、CH4、铜元素等)表现出强相关性,进一步证明了地球深部发生的宏观尺度上的构造活动对热泉流体栖息微生物的种群结构影响极大(图5)。
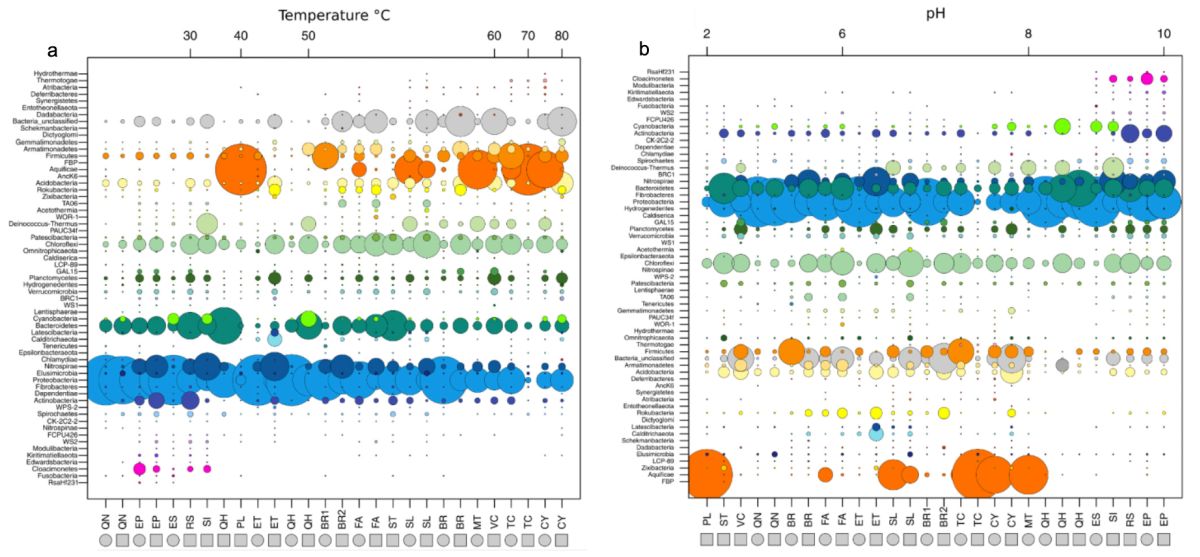
图3 热泉流体中栖息的微生物类群(门水平)及其与温度(a)和pH(b)的相关性(见Fullerton et al., 2021补充材料)
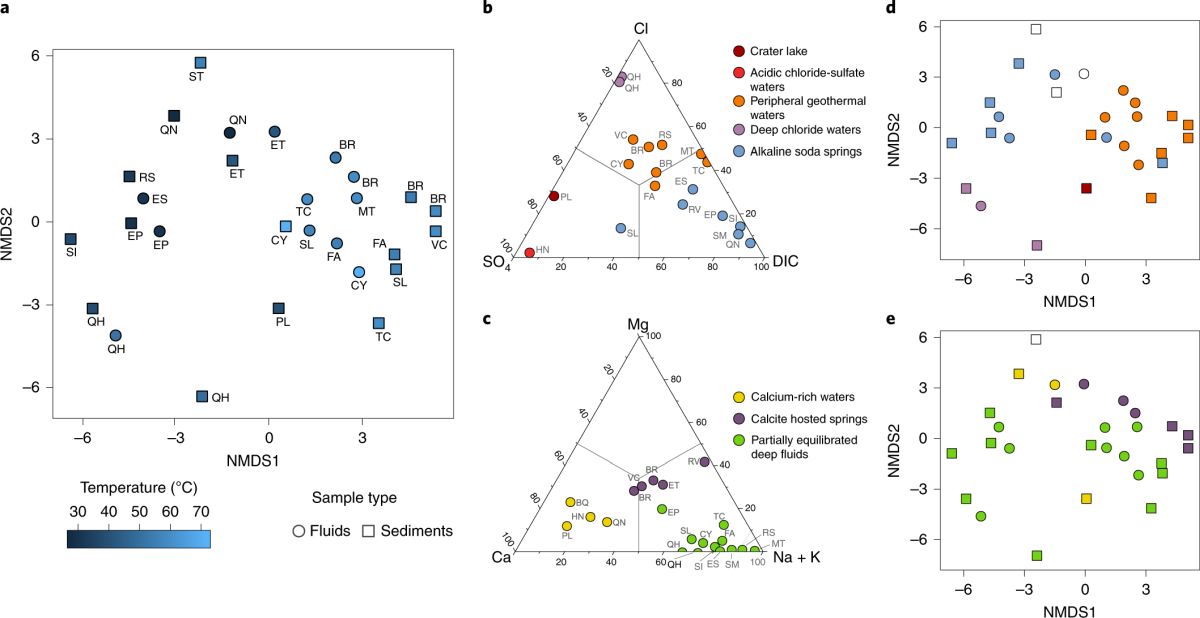
图4 微生物的种类与热泉流体中阴阳离子组分的相关性(Fullerton et al., 2021)。 (a)微生物多样性与温度的关系;(b)热泉中基于主要阴离子的聚类三元图;(c)热泉中基于主要阳离子的聚类三元图; (d)微生物多样性与阴离子的相关性;(e)微生物多样性与阳离子的相关性。注:d和e中的样品名与a中的一致
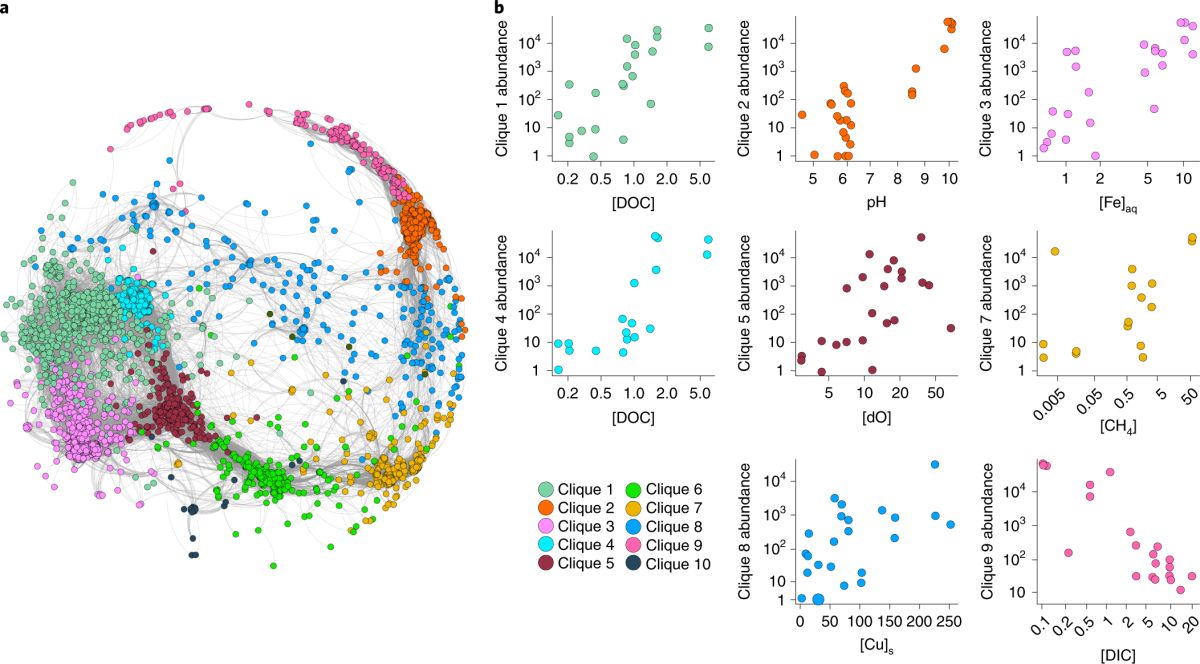
图5 微生物的种类与俯冲带地球化学环境因子之间的相关性(Fullerton et al., 2021)。 (a)所有样品的主要 ASVs 的共现网络分析; (b)不同微生物群落中ASVs与相应的环境因子之间的关系
为了进一步寻求化能自养微生物与碳的地球化学循环之间联系的基因方面的直接证据,该研究还利用宏基因组的中心碳代谢途径相关的功能基因进行共现网络分析(图6a)。热泉流体微生物的宏基因组中主要发现了编码以下三种固碳途径关键酶的功能基因,即Wood–Ljungdahl、rTCA和Calvin–Benson–Bassham (图6a)。其中rTCA循环相关的基因丰度 与 DIC浓度呈现出很好的的相关性,表明DIC和基于rTCA固碳途径的化能自养微生物之间存在密切关系(图6b)。
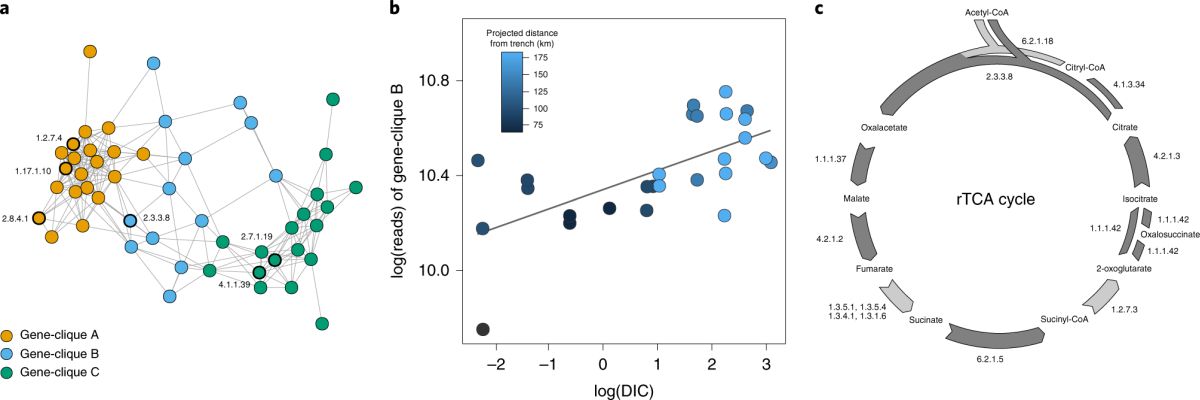
图6 微生物固碳功能基因的共现网络分析及其与环境参数的相关性(Fullerton et al., 2021)。(a) 中心碳代谢途径功能基因的共现网络分析。橙色代表Wood-Ljungdahl途径,蓝色代表 rTCA途径,绿色代表Calvin-Benson-Bassham途径;(b) rTCA途径功能基因丰度(reads数)与DIC浓度的正相关性;(c) rTCA固碳途径涉及的关键酶
基于前面的分析可以看出,发生于宏观尺度的地球深部构造活动对地球深部生物圈的生物量及其多样性均产生着重要的影响。此外,基于对微生物细胞周转期和碳含量的估计,研究人员发现微生物化学自养过程生成整个DOC储库仅需要44-442年,而该区深部流体的滞留时间长达8.8×104-8.8×106年,对于整个DOC储库的形成而言时间是足够的。而且,与地球深部贫营养的地下含水层中化能自养的生态系统相比,汇聚板块边缘的化能自养系统能将DOC的浓度提升100倍,达到毫摩尔级别。基于生命能够忍受的温度上限为122°C的等温线,估算在哥斯达尼加北部汇聚板块边缘地表一下岩石深部生物圈的生物碳含量为7.9×1010至7.9×1011 mol,即使只有30%的地下深部微生物属于化能自养型微生物,它们对俯冲板片产生的CO2的年均生物固碳量也将达到1.4 ×109至1.4 ×1010 mol,这一生物固碳量比单独计算DOC的碳量要大得多(4.1 ×105 mol至 4.4 ×108 mol),从而使进入俯冲带的碳量额外减少了1%-13%,并使进入深部地幔的碳量进一步减少了2%到22%。化能自养微生物的固碳作用是方解石、DIC和DOC碳封存的一个重要补充。前面基于宏基因组的研究结果表明,这些化石自养生物受到DIC可用性的限制(图6b),因为方解石沉淀是弧前区碳汇的主要方式,消耗了大量的DIC,所以化能自养微生物可利用的 DIC受限,进而影响到生物的固碳效率。但是,在其它缺乏方解石碳封存能力的汇聚板块边缘,化能自养微生物将能够获得充足的DIC,这将大大提高微生物固碳的效率,从而有效弥补区域的碳封存能力。因此,微观的深部化能自养微生物固碳活动是对板块俯冲区方解石、DIC和DOC等其它常规碳封存方式的有效补充,从而影响了宏观的地球深部活动。
综上所述,哥斯达黎加汇聚板块边缘栖息的微生物群落结构与来自地球深部构造过程的地球化学信号密切相关。岩石深部生物圈微生物群落的多样性随着汇聚板块边缘所提供的氧化还原活性底物以及DIC供应的变化而不同,而从下沉的板片和地幔中流动的挥发物和元素在返回地表的过程中通过与深部生物圈的相互作用而发生改变,形成汇聚板块边缘地质圈和生物之间的反馈耦合,从而影响汇聚板块边缘的挥发物的通量。这对理解深时碳储库的变化、地幔碳封存与地壳碳封存的耦合以及最终的行星氧化还原平衡和气候长期稳定性具有重要意义。
主要参考文献
Fullerton K M, Schrenk M O, Yücel M, et al. Effect of tectonic processes on biosphere–geosphere feedbacks across a convergent margin[J]. Nature Geoscience, 2021, 14(5): 301-306.(原文链接)
Power J F, Carere C R, Lee C K, Wakerley G L J, Evans D W, Button M, White D, Climo M D, Hinze A M, Morgan X C, McDonald I R, Cary S C, Stott M B. Microbial biogeography of 925 geothermal springs in New Zealand[J]. Nature Communications, 2018, 9(1): 2876.
Schellart W P, Rawlinson N. Convergent plate margin dynamics: New perspectives from structural geology, geophysics and geodynamic modelling[J]. Tectonophysics, 2010, 483(1-2): 4-19.
张传伦, 林间, 李三忠等. 构造微生物学:地球生物学研究的新理念[J]. 中国科学:地球科学, 2018, 48(7): 956-959
(撰稿:刘嘉玮/南洋理工大学数理学院,李江涛/同济大学海洋与地球科学学院,李金华/地质地球所地星室)