在寒武纪中期至奥陶纪时期,曾发生过地球生命历史中最显著的生物多样性变化:奥陶纪生物大辐射事件(GOBE)和晚奥陶世生物灭绝事件。这些事件是在一个动荡的地球系统背景下发生的,致使学者们提出了多种关于生物多样性变化与寒武-奥陶纪地球系统演变之间因果联系的假说。例如,GOBE事件是对气候变冷和随之生理压力减缓的响应(Trotter et al., 2008; Rasmussen et al., 2016),而晚奥陶世大灭绝则与冈瓦纳冰期有关,后者可通过变冷与海面下降的共同作用(Finnegan et al., 2011)或者海洋通气能力的变化(Hammarlund et al., 2012)来影响生物。气候变化与造山运动、岛弧增生和蛇绿岩逆冲作用的增强有关,它们通过火山或者硅酸盐岩风化过程能影响大气CO2浓度(Buggisch et al., 2010; Swanson-Hysell and Macdonald, 2017; Macdonald et al., 2019)。
验证构造、气候和生物演化相互作用的假说需要可靠的记录。化石提供了早显生宙以来丰富的海洋生物多样性记录,构造变动历史也能通过多维地质证据得到重建。然而,早古生代气候变化却缺乏很好的约束,主要是因为:一、传统氧同位素(δ18O)温度计重建古温度有很大不确定性(因为缺少独立的海水氧同位素数据);二、寒武纪和早奥陶世沉积岩中缺乏保存完好的海洋生物(如腕足类和牙形石)壳体化石(即重建古气候的理想载体)。
碳酸盐团簇同位素技术的出现,解决了如上所述的第一个难题。已有数据表明,早古生代生物碳酸盐δ18O值系统地低于现代海洋碳酸盐。这两个时代的同位素差异有三种可能解释:早古生代海水温度比现代高、早古生代海水δ18O值比现代低、成岩作用导致早古生代碳酸盐δ18O变低。然而,生物壳体化石的团簇同位素研究表明,除了受冰量效应和局地蒸发富集的影响,古生代海水氧同位素组成与新生代无冰时的海洋相似(Bergmann et al., 2018)。由此,早古生代较低δ18O值是气候变化、成岩作用、蒸发富集和冰量效应等共同作用的结果。
最近,美国麻省理工大学的Goldberg博士和合作者为了弥补化石材料缺失的不足,对全球7个地区(多数在热带,见图1)的寒武纪-奥陶纪碳酸盐岩台地剖面中沉积物全岩稳定同位素进行系统地分析、收集和集成研究,建立了全球首条高分辨率早古生代海洋δ18O曲线,相关成果发表于PNAS。
图1 本文中各研究地点的古地理位置(以465 Ma为例)(Goldberg et al., 2021)
他们首先对位于挪威斯瓦尔巴群岛和加拿大纽芬兰岛的早古生代碳酸盐岩台地中保存较好部位的微晶碳酸盐进行了团簇同位素和碳、氧同位素分析,来揭示成岩作用的发生机制。结果表明,团簇同位素温度为45~100℃,受到成岩作用改造而发生了重置。但是,两个研究地点的样品在团簇同位素温度、碳酸盐矿物δ18O值和同位素平衡流体δ18O值构成的平面图中却具有相似的地球化学轨迹(图2),说明他们所经历的成岩改造发生在相对封闭的体系(即具有较低的水/岩比)。在这种情况下,随着成岩作用进行,流体同位素组成会朝着岩石同位素值方向演化,由此可以解释流体中相对富集的同位素组成(0~11‰ VSMOW),以及温度和流体同位素组成沿着碳酸盐矿物δ18O等值线方向展布的相关关系(图2)。由此,埋藏过程中流体较少而使得碳酸盐矿物氧同位素组成被极大地保留了下来。
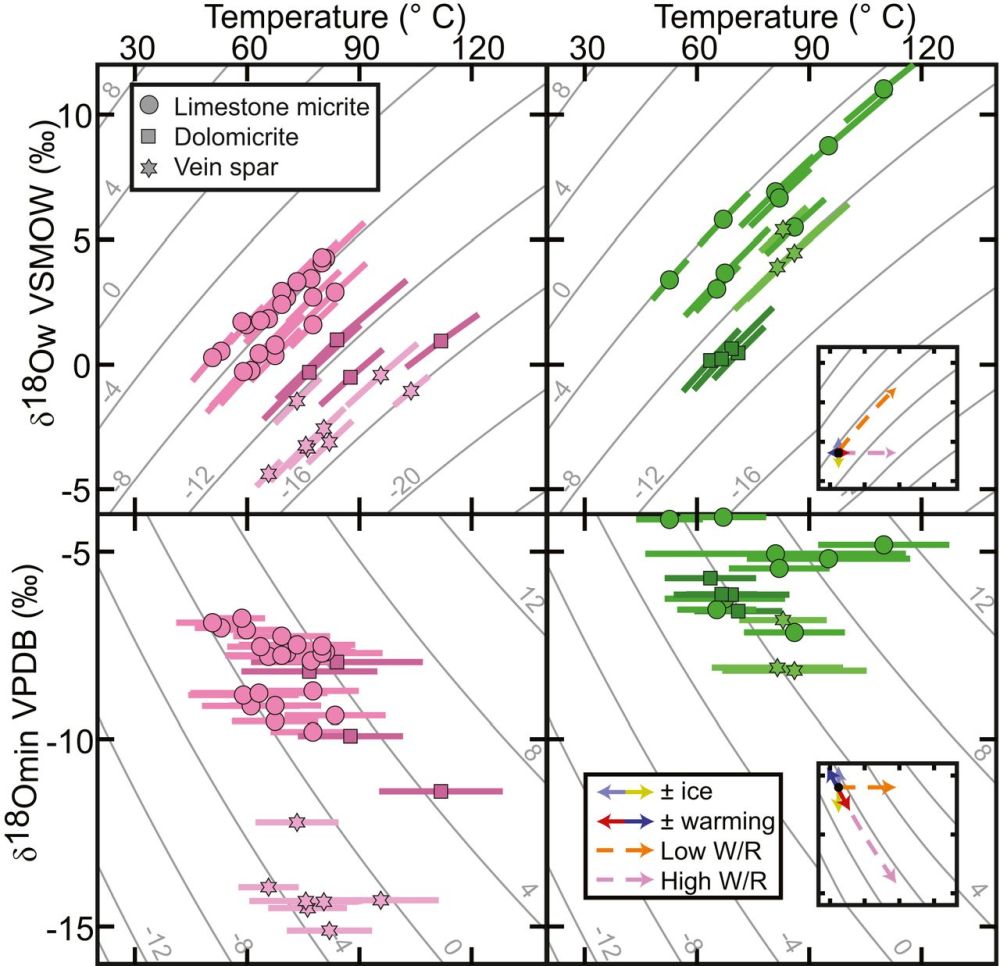
图2 来自斯瓦尔巴群岛(左)和纽芬兰岛(右)样品的团簇同位素温度、碳酸盐矿物δ18O和流体δ18O之间的对比。插图显示地表条件(实线)和成岩作用方式(虚线)的理论变化轨迹。各图中灰色线条表示方解石平衡分馏变量的等值线。W/R为水/岩比值(Goldberg et al., 2021)
在上述结果的激励下,他们收集了其他研究区域已发表的寒武纪-奥陶纪碳酸盐岩台地全岩δ18O数据。为了减少局地效应和成岩作用的影响,他们将区域δ18O记录进行平均化处理,最终集成了一条全球热带浅海δ18O曲线。该集成的全岩氧同位素记录与腕足类和牙形石δ18O数据在变化趋势上具有很好的一致性(图3),显示了数据的可靠性。
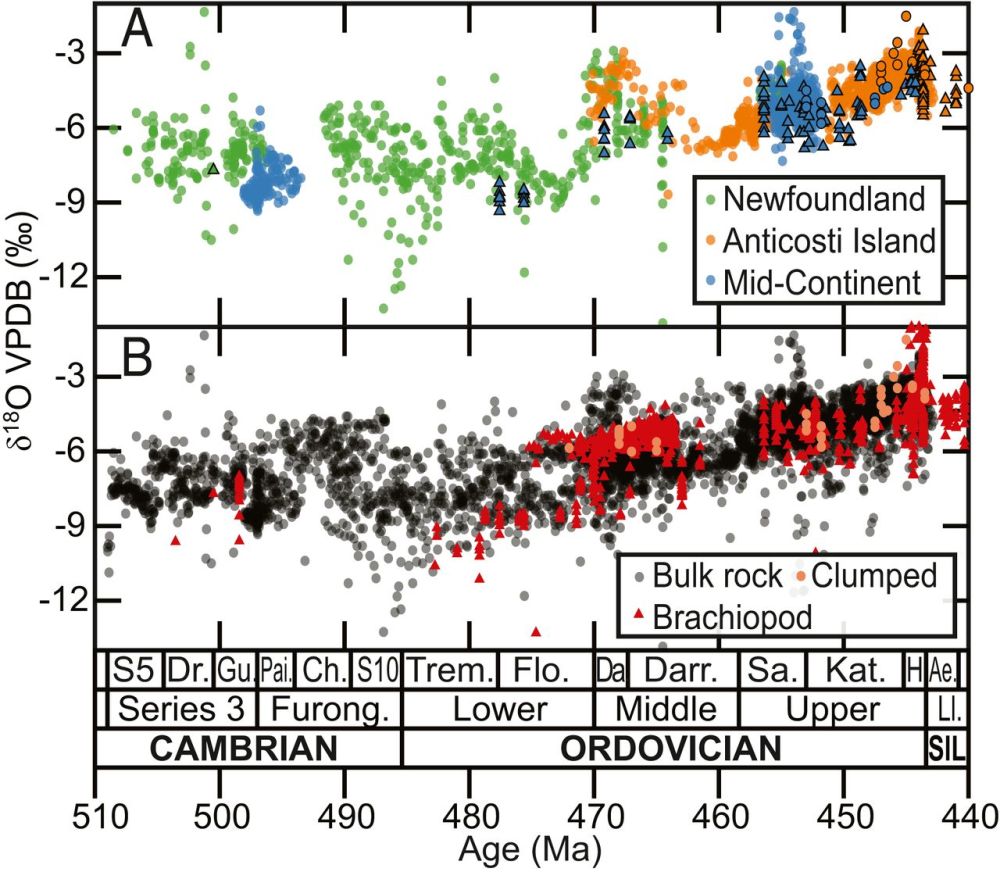
图3 全岩与壳体化石氧同位素数据对比。(A)来自北美样品的数据。圆形代表全岩氧同位素,三角形代表腕足类壳体氧同位素。(B)来自全球样品的数据,包括全岩(黑色圆点)、腕足类壳体(红色三角形)和团簇同位素样品δ18O(橙色圆点)(Goldberg et al., 2021)
基于前人已发表保存完好的腕足类和珊瑚等化石团簇同位素温度和氧同位素数据,作者采用模型计算得到当时海水氧同位素组成,比如奥陶纪大部分时段海水δ18O值为-1.4‰ VSMOW,仅在晚奥陶世冰期由于冰量驱动有所增加。根据重建的海水δ18O变化和碳酸盐岩全岩δ18O值,Goldberg等人计算获得了寒武纪-奥陶纪地表温度变化记录(图4)。由于成岩作用通常会使δ18O值变低(趋向更高的重建温度),所以Goldberg等人认为重建温度记录中最低十分位数部分才最能代表热带浅海温度,而最高十分位数部分可能反映了成岩改造。该最低十分位数部分的古温度数据表明,早奥陶世(特马豆克阶-弗洛阶)温度显著高于寒武纪晚期和中-晚奥陶世时期。在该温暖期之后,弗洛阶-大坪阶时段以约1℃/Ma的速率降温10~15℃。尽管在距今467 Ma之后晚奥陶世温度估算对模拟的冰盖增长比较敏感,本研究记录重建的温度却显示454~449 Ma时段气候温暖,随后才进入较冷的赫南特冰室状态(图4B)。

图4 早期动物演化的气候和构造背景,演化事件被突出显示。(A)全岩δ13C值,主要同位素漂移事件已被标注。GICE为古登堡δ13C漂移,HICE为赫南特δ13C漂移,而SPICE则为排碧阶碳同位素正漂移。(B)氧同位素温度,用不同明暗度来表示全球所有数据的十分位数分布。暗色区域代表数据中18O更富集(温度更低)的部分,具有较弱的成岩作用改造。红色圆点为腕足类氧同位素温度,蓝色菱形为牙形石氧同位素温度。(C)活动构造缝合带长度。黑色线为总活动长度,彩色线代表热带地区不同纬度的活动长度。(D)锶同位素比值,为大陆风化替代指标。(E)冰盖的纬向范围。(F)绿色曲线(区域)代表海洋种属多样性(左侧坐标轴),紫色虚线代表含有叠层石的海相碳酸盐比例(右侧坐标轴)。纵向灰色条带为GOBE发生时段,竖线指示广泛分布的K-斑脱岩年龄
本研究高分辨率温度记录使得作者能够评估构造、气候和生物演化之间相互作用(图4)。寒武纪/奥陶纪界限和早奥陶世时期的高温可能是当时物种调整速率增加的一个主要驱动因素,而晚寒武世和早奥陶世浅海大范围缺氧(水温与溶解氧的反向关系)又潜在地加剧了该生物事件的发生。前人研究认为跨越早/中奥陶世界限的变冷增加了海洋含氧量,从而可能触发了GOBE事件(Trotter et al., 2008)。然而,本研究表明仅有中奥陶世较短时间段内变冷才与波罗的海和劳伦古陆发生的GOBE主要脉动相对应(图4B和4F)。同时,碳酸盐δ13C的增加也与变冷有关,主要表现在中-晚奥陶世长期变化趋势和与赫南特冰期相关的碳同位素大幅正漂(图4A)。叠层石的丰度呈现出与温度相同、与动物多样性相反的变化趋势(图4F),表现为晚寒武世丰度增加和早-中奥陶世界限时期丰度降低。上述温度与生物多样性的相关关系暗示了气候及其相关因子(如海水氧化程度)对微生物和后生动物相对丰度和多样性的影响。
构造运动是早古生代气候的最终驱动因素,进而影响生物多样性变化。例如,中奥陶世温度变冷与岛弧增生的加强是同时的(图4C),这与前人提出的变冷的构造驱动模型(Macdonald et al., 2019)是一致的,但却超前于87Sr/86Sr值的降低(该滞后可能是由于海洋中Sr的滞留时间长)(图4C)。晚奥陶世中穿插的短暂温暖期与反复发生的区域火山事件有关,这被广泛分布的定年较好的K-斑脱岩火山灰层和汞化学地层所证实(Jones et al., 2017; Yang et al., 2019)。火山活动既可通过CO2直接排放而导致变暖,又可以通过硅酸盐岩风化吸收CO2而导致变冷,从而导致气候的不稳定性。该时期气候变率的增加与GOBE事件结束同步,自此开始了长达8百万年之久生物多样性降低(在赫南特冰期最盛期达到最低)和生物灭绝。
本文采用团簇同位素手段揭示了成岩作用对早古生代碳酸盐δ18O影响很小,然后通过全球集成的全岩δ18O数据首次获得了寒武纪-奥陶纪高分辨率气候记录,并将该记录置于生物演化和构造变动的背景下进行对比研究,明确指出地球气候与构造和火山活动关系紧密,对早期动物演化中生物多样性的变化具有较强的控制作用。Goldberg等人所展示的研究方法为我们解释其他深时阶段(如前寒武纪)全岩δ18O记录提供了很好的借鉴。
主要参考文献
Bergmann K D, Finnegan S, Creel R, et al. A paired apatite and calcite clumped isotope thermometry approach to estimating Cambro-Ordovician seawater temperatures and isotopic composition[J]. Geochemica et Cosmochimica Acta, 2018, 224: 18-41.
Buggisch W, Joachimski M M, Lehnert O, et al. Did intense volcanism trigger the first Late Ordovician icehouse?[J]. Geology, 2010, 38: 327-330.
Finnegan S, Bergmann K, Eiler J M, et al. The magnitude and duration of Late Ordovician–Early Silurian glaciation[J]. Science, 2011, 331: 903-906.
Goldberg S L, Present T M, Finnegan S, et al. A high-resolution record of early Paleozoic climate. Proceedings of the National Academy of Sciences, 2021, 118: e2013083118.(原文链接)
Hammarlund E U, Dahl T W, Harper D A T, et al. A sulfidic driver for the end-Ordovician mass extinction[J]. Earth Planet Science Letters, 2012, 331/332: 128-139.
Jones D S, Martini A M, Fike D A, et al. A volcanic trigger for the late Ordovician mass extinction? Mercury data from south China and Laurentia[J]. Geology, 2017, 45: 631-634.
Macdonald F A, Swanson-Hysell N L, Park Y, et al., Arc-continent collisions in the tropics set Earth’s climate state[J]. Science, 2019, 364: 181-184.
Rasmussen C M ?, Ullmann C V, Jakobsen K G, et al. Onset of main Phanerozoic marine radiation sparked by emerging mid Ordovician icehouse[J]. Scientific Reports, 2016, 6: 18884.
Swanson-Hysell N L, Macdonald F A. Tropical weathering of the Taconic orogeny as a driver for Ordovician cooling[J]. Geology, 2017, 45: 719-722.
Trotter J A, Williams I S, Barnes C R, et al. Did cooling oceans trigger Ordovician biodiversification? Evidence from conodont thermometry[J]. Science, 2008, 321: 550-554.
Yang S, Hu W S, Wang X L, et al. Duration, evolution, and implications of volcanic activity across the Ordovician–Silurian transition in the Lower Yangtze region, South China[J]. Earth Planet Science Letters, 2019, 518: 13-25.
(撰稿:王旭/新生代室)